© Borgis - Postępy Fitoterapii 1/2006, s. 36-40
*Justyna Stefanowicz-Hajduk, J. Renata Ochocka
Saponiny steroidowe – występowanie, właściwości i zastosowanie w lecznictwie
Steroidal saponins – occurrence, characteristic and application in therapeutics
Katedra i Zakład Biologii i Botaniki Farmaceutycznej Akademii Medycznej w Gdańsku
Kierownik Katedry i Zakładu: prof. dr hab. n. farm. J. Renata Ochocka
Summary
Steroid saponins are glycosides distributed in many species of plants widely, mainly in the monocotyledons order ( Monocotyledones). They consist of sugar units and an aglycone known as a "sapogenin”, which has a steroid structure. Saponins have application in industry mainly as precursors for corticosteroids and hormonal compounds derivatives of pregnane due to the character of their steroid molecule.

Saponiny (saponozydy) to grupa związków glikozydowych wykazujących szereg wspólnych właściwości, a rozpowszechnionych w ponad 85 rodzinach systematycznych. Występują głównie u jednoliściennych ( Monocotyledones) w rodzinach: Dioscoreaceae (pochrzynowate), Liliaceae (liliowate), Agavaceae (agawowate), rzadziej natomiast u dwuliściennych ( Dicotyledones) w: Scrophulariaceae (trędownikowate), Fabaceae (motylkowate), Solanaceae (psiankowate), Ranunculaceae (jaskrowate), Asteraceae (złożone) (10).
Saponiny są bezazotowymi związkami organicznymi nie spotykanymi w świecie zwierzęcym. Składają się z części cukrowej (łańcucha 3-5 jednocukrów) oraz aglikonu – sapogeniny, mającego charakter sterolowy bądź trójterpenowy. Charakter trójterpenowy sapogeniny uwarunkowany jest najczęściej obecnością aglikonu typu α-amyryny o 30 atomach węgla, natomiast charakter sterolowy obecnością zasadniczego rdzenia steranu o 27 atomach węgla z bocznym ugrupowaniem cyklicznym w pozycji C17 (ryc. 1). Wśród saponin steroidowych wyróżnia się dodatkowo pochodne spirostanu oraz furostanu (ryc. 2). Poszczególne saponiny – steroidowe i trójterpenowe różnią się liczbą i położeniem grup funkcyjnych, ilością i miejscem występowania podwójnych wiązań oraz rodzajem i ilością cukrów (16, 19).
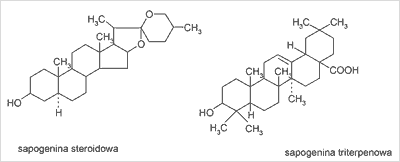
Ryc. 1. Wzór sapogeniny steroidowej oraz triterpenowej.
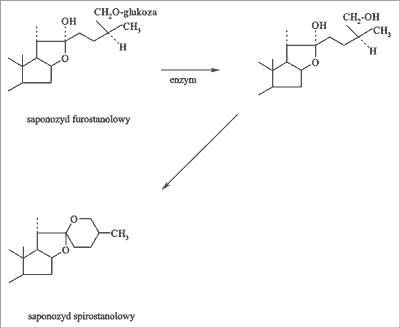
Ryc. 2. Przebieg procesu powstawania saponiny spirostanolowej.
W części cukrowej wszystkich saponin występuje przeważnie glukoza, galaktoza, ramnoza, ksyloza, fukoza, arabinoza, rzadziej kwas galakturonowy i glukuronowy. Łańcuch cukrowy może być prosty bądź rozgałęziony, mogą też występować dwa lub trzy oddzielne łańcuchy cukrowe, stąd też saponiny o jednym łańcuchu nazwano monodesmozydami, o dwóch – bidesmozydami, a o trzech – tridesmozydami (18).
Saponiny obniżają napięcie powierzchniowe pomiędzy heterogennymi układami, mają właściwości hemolityczne oraz łączą się z cholesterolem. Wspomniane obniżenie napięcia powierzchniowego w roztworach wodnych przejawia się efektem pienienia. Na tej reakcji oparte jest przybliżone oznaczanie ilości saponin w wyciągach przez wyznaczenie tzw. wskaźnika (indeksu) pienienia się. Inną metodą oznaczenia saponin jest wyznaczenie wskaźnika hemolitycznego, tj. najwyższego rozcieńczenia danej saponiny, przy którym powoduje ona jeszcze całkowitą hemolizę określonej ilości krwinek baranich lub bydlęcych.
Ponadto większość saponin należy do silnych trucizn dla zwierząt zimnokrwistych. Na tej właściwości oparte jest oznaczanie wskaźnika toksyczności saponin, tj. oznaczanie krańcowe stężenia zabijającego rybki w ciągu godziny (16, 18).
Saponiny steroidowe występują w wielu roślinach, jednak większe znaczenie w przemyśle farmaceutycznym mają te gatunki, które stanowią źródło surowców wyjściowych do syntezy kortykosteroidów i hormonów płciowych.
Zainteresowanie saponinami wzrosło w XX wieku, kiedy to podjęto liczne badania nad ich strukturą i zastosowaniem jako prekursorów kortyzonu – hormonu pierwotnie wyizolowanego z kory nadnerczy i w późniejszym okresie również zsyntezowanego z kwasów żółciowych. Poszukiwania roślinnych prekursorów opierały się przede wszystkim na tym, czy wspomniany prekursor posiada grupy hydroksylowe w 3- i 11- pozycji w cząsteczce, bądź też czy istnieje możliwość łatwego przekształcenia związku do wspomnianych struktur. Stąd najbardziej obiecującymi saponinami okazały się: diosgenina i botogenina występujące w roślinach rodzaju Dioscorea (pochrzyn, ignam, jams), hekogenina, manogenina oraz gitogenina z gatunków rodzaju Agave (agawa), sarsasapogenina i smilagenina z rodzaju Smilax (kolcorośl, sarsaparyla), sarmentogenina z rodzaju Strophanthus (strofant).
W pozyskiwaniu wymienionych związków największe znaczenie wśród jednoliściennych mają rodziny Liliaceae, Amaryllidaceae i Dioscoreaceae, natomiast wśród dwuliściennych za najbardziej obiecujący był początkowo uznawany rodzaj Strophanthus z rodziny Apocynaceae. Większość jednak prac dotyczących wykorzystania roślinnych metabolitów w syntezie kortyzonu koncentrowała się na trzech wymienionych rodzinach z klasy jednoliściennych (20, 21). Szczególnym zainteresowaniem cieszyły się gatunki rodzaju Dioscorea, a to ze względu na obecność diosgeniny, która jest jednym z głównych związków służących do otrzymywania glikokortykosteroidów, z zastosowaniem transformacji mikrobiologicznej.
Obecnie podstawowym źródłem otrzymywania rdzenia steroidowego stosowanego do syntezy hormonów są rośliny, podczas gdy do niedawna głównym źródłem otrzymywania hormonów steroidowych były gonady oraz gruczoły nadnerczy zwierząt. Ilość hormonów uzyskanych z narządów zwierzęcych była bardzo niewielka przy stosunkowo dużej ilości pobranych gruczołów (w 1934 roku Schering Laboratories w Berlinie otrzymał zaledwie 20 mg czystego krystalicznego progesteronu z 625 kg jajników uzyskanych z 50 tys. świń) (21).
Jednym z pionierów w dziedzinie otrzymywania hormonów steroidowych z naturalnie występujących związków jest Russell E. Marker, który opracował proces przemiany sapogenin steroidowych do progesteronu (w 1943 roku wyizolował diosgeninę z Dioscorea mexicana i otrzymał 3 kg progesteronu) (ryc. 3). Kwestia przekształcenia i otrzymania z progesteronu 11-α-hydroksyprogesteronu została rozwiązana w latach 50-XX wieku, kiedy to odkryto, że mikroorganizm Rhizopus arrhizus potrafi zamienić wspomniany progesteron w hydroksyprogesteron z wydajnością 80-90% (ryc. 4) (21). Okazało się również, że do produkcji hormonów płciowych nadają się tylko sapogeniny mające jedną grupę hydroksylową w pozycji 3, a w celu otrzymania kortykosteroidów (kortyzonu) konieczne jest wprowadzenie grupy hydroksylowej w pozycję 11 (10). Można to przeprowadzić, jak już wspomniano wcześniej, mikrobiologicznie. Najlepszymi byłyby oczywiście w tym przypadku naturalne związki steroidowe, posiadające już grupę OH w pozycji 11, jednak takich naturalnych związków jest niewiele (większe znaczenie ma jedynie sarmentogenina, która występuje w nasionach Strophanthus sarmentosus, jest jednak trudno dostępna).
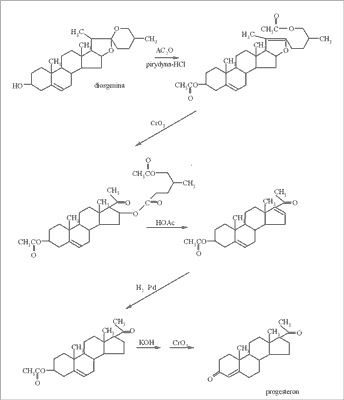
Ryc. 3. Schemat przemiany diosgeniny w progesteron wg Markera.
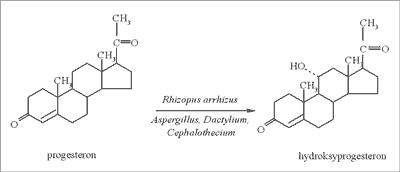
Ryc. 4. Schemat przemiany progesteronu w 11α – hydroksyprogesteron.
Powyżej zamieściliśmy fragment artykułu, do którego możesz uzyskać pełny dostęp.
Mam kod dostępu
- Aby uzyskać płatny dostęp do pełnej treści powyższego artykułu albo wszystkich artykułów (w zależności od wybranej opcji), należy wprowadzić kod.
- Wprowadzając kod, akceptują Państwo treść Regulaminu oraz potwierdzają zapoznanie się z nim.
- Aby kupić kod proszę skorzystać z jednej z poniższych opcji.
Opcja #1
24 zł
Wybieram
- dostęp do tego artykułu
- dostęp na 7 dni
uzyskany kod musi być wprowadzony na stronie artykułu, do którego został wykupiony
Opcja #2
59 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 30 dni
- najpopularniejsza opcja
Opcja #3
119 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 90 dni
- oszczędzasz 28 zł
Piśmiennictwo
1. Applezweig N. Steroids. Chem Week. 1969, 104, 57. 2. Corbiere C., Liagre B., Bianchi A., Bordji K., Dauca M., Netter P., Beneytout J.L. Different contribution of apoptosis to the antiproliferative effects of diosgenin and other plant steroids, hecogenin and tigogenin, on human 1547 osteosarcoma cells.Int J Oncol. 2003, 22(4), 899. 3. Corbiere C., Liagre B., Terro F., Beneytout J.L. Induction of antiproliferative effect by diosgenin through activation of p53, release of apoptoso – inducing factor (AIF) and modulation of caspase – 3 activity in different human cancer cells. Cell Res. 2004, 14(3), 188. 4. Evans WC. Advances in pharmacognosy. Manuf Chem Aerosol News. 1956, 27, 51. 5. Fazli FR. Contraceptives and other steroid drugs: their production from steroidal sapogenins. Pak J Sci. 1968, 20(1, 2), 64. 6. Furmanowa M., Guzewska J., Bełdowska B., Rzędowski M. Gatunki zawierające diosgeninę w kulturze tkankowej in vitro I. Wpływ warunków hodowli na wzrost tkanek Trigonella foenum – graecum L. i zawartość diosgeniny. Herba Polon. 1985, 31, 3. 7. Harborne J.B., Baxter H., Moss G.P. Phytochemical dictionary: a handbook of bioactive compounds from plants. Taylor and Francis, London 1999. 8. Huo R., Zhou Q., Wang B., Tashiro S., Onodera S., Ikejima T. Diosgenin induces apoptosis in HeLa cells via activation of caspase pathway. Acta Pharmacol Sin. 2004, 25(8), 1077. 9. Juarez – Oropeza MA., Diaz – Zagoya JC., Rabinowitz JC. In vivo and in vitro studies of hypocholesterolemic effects of diosgenin in rats. Int J Biochem. 1987, 19(8), 679. 10. Kohlmünzer S. Farmakognozja. Wyd. Lekarskie PZWL, Warszawa 1998. 11. Kokot F. Choroby wewnętrzne. Wyd. Lekarskie PZWL, Warszawa 1996. 12. Liagre B., Vergne-Salle P., Corbiere C., Charissoux J.L., Beneytout J.L. Diosgenin, a plant steroid, induces apoptosis in human rheumatoid arthritis synoviocytes with cyclooxygenase-2 overexpression. Arthritis Res Ther. 2004, 6, R373. 13. Liu MJ., Wang Z., Ju Y., Wong RN., Wu QY. Diosgenin induces cell cycle arrest and apoptosis in human leukemia K562 cells with the disruption of Ca2+ homeostasis. Cancer Chemother Pharmacol. 2004, 55(1), 79. 14. Matławska I. Leki roślinne w terapii guzków krwawniczych odbytu. Postępy Fitoterapii. 2002, 3-4. 15. Moalic S., Liagre B., Corbiére C., Bianchi A., Dauça M., Bordji K., Beneytout J.L. A plant steroid, diosgenin, induces apoptosis, cell cycle arrest and COX activity in osteosarcoma cells. FEBS Lett. 2001, 506, 225. 16. Muszyński J. Farmakognozja. Wyd. Lekarskie PZWL, Warszawa 1957. 17. Oncina R., Botia J.M., Del Rio J.A., Ortuno A. Bioproduction of diosgenin in callus cultures of Trigonella foenum – graecum L. Food Chem. 2000, 70, 489. 18. Osbourn A. Saponins and plant defence – a soap story. Trends Plant Sci. 1996, 1, 1. 19. Strzelecka H., Kamińska J., Kowalski J., Walewska E. Chemiczne metody badań roślinnych surowców leczniczych. Wyd. Lekarskie PZWL, Warszawa 1982. 20. Strzelecka H., Kowalski J. Encyklopedia zielarstwa i ziołolecznictwa. Wyd. Naukowe PWN, Warszawa 2000. 21. Tyler V.E., Brady L.R., Robbers J.E. Pharmacognosy. Lea&Febiger, Philadelphia 1988. 22. Yokosuka A., Mimaki Y., Kuroda M., Sashida Y. A New steroidal saponin from the leaves of Agave americana. Planta Med. 2000, 66(4), 393.