© Borgis - Postępy Nauk Medycznych 11/2011, s. 917-923
*Marlena Godlewska1, Andrzej Gardas1, Monika Góra2
Biosynthesis, post-translational modifications, maturation, and physiological function of human thyroid peroxidase**
Biosynteza, modyfikacje postranslacyjne, dojrzewanie i fizjologiczna funkcja ludzkiej peroksydazy tarczycowej
1Department of Biochemistry and Molecular Biology, Medical Centre of Postgraduate Education, Warsaw
Head of Department: prof. dr hab. Barbara Czarnocka
2Department of Genetics, Institute of Biochemistry and Biophysics Polish Academy of Sciences, Warsaw
Head of Department: prof. dr hab. Teresa Żołądek
Streszczenie
Peroksydaza tarczycowa (TPO) jest jednocześnie głównym autoantygenem w autoimmunologicznych chorobach tarczycy i kluczowym enzymem, wraz z białkami DUOX, odpowiedzialnym za syntezę hormonów tarczycy. Jeszcze na poziomie retikulum endoplazmatycznego nowo zsyntetyzowana TPO przechodzi proces N-glikozylacji i włączania hemu oraz przejściowo oddziałuje z białkami opiekuńczymi (chaperonami). Następnie podczas transportu przez aparat Golgiego do cząsteczki TPO dołączane są O-glikany, natomiast oligosacharydy połączone z białkiem wiązaniem N-glikozydowym poddawane są dalszym modyfikacjom. Po opuszczeniu aparatu Golgiego, ale przed dotarciem na powierzchnię komórki, z peroksydazy zostaje usunięty propeptyd. TPO tworzy dimery, jednak mechanizm tego procesu jest nadal nieznany. Tylko część zsyntetyzowanego białka dociera do błony komórkowej, natomiast cząsteczki z częściowym lub całkowitym zaburzeniem struktury przestrzennej są degradowane w retikulum endoplazmatycznym przez odpowiednio proteasom lub proteazę podobną do plazminogenu.
Summary
Apart from being the major target autoantigen in autoimmune thyroid disease, thyroid peroxidase (TPO) is the key enzyme, together with DUOX proteins, responsible for the thyroid hormone generation. Newly synthesized TPO is processed in the endoplasmic reticulum, undergoing N-glycosylation, heme fixation, and transient interaction with molecular chaperones. Then, while transporting through the Golgi apparatus, O-glycans are attached to the TPO molecule, whereas N-linked oligosaccharides undergo further modifications. After exiting the Golgi, but before it reaches the cell surface, the propeptide is removed. TPO forms dimers, however, the mechanism of this process is still unknown. Only part of the synthesized TPO reaches the cell membrane, whereas partially or totally misfolded forms of TPO are degraded in the endoplasmic reticulum by the proteasome or a plasminogen-like protease, respectively.

Introduction
Since 1985 we have known that human thyroid peroxidase (TPO), previously known as the thyroid microsomal antigen, is a target for the autoimmune response in autoimmune thyroid diseases (1). The majority of patients’ autoantibodies to thyroid peroxidase are directed to two discontinuous determinants on the protein surface. Several approaches have been taken by our and other laboratories to localize the autoantibody binding regions on TPO. These studies provided a greater insight into the nature of the autoimmune response to the thyroid. In a few recent reviews the current knowledge on TPO as an autoantigen was summarized (2-5). Here, we focus on the biochemical aspects of thyroid peroxidase, especially on its maturation process, trafficking and participation in thyroid hormone biosynthesis.
Protein structure and expression
The human TPO gene, that was cloned in 1987 (6-8), is located on chromosome 2 (region pter-p12) and it is composed of 17 exons and 16 introns (9). The TPO expression is controlled by thyroid-specific factors, namely TTF-1, TTF-2, and Pax-8 (reviewed in Ref. 10-12). Till now several isoforms of TPO cDNA were identified, cloned and characterized (reviewed in Ref. 2, 10).
TPO belongs to the animal peroxidase protein family together with myeloperoxidase (MPO), lactoperoxidase (LPO), eosinophil peroxidase (EPO) (reviewed in Ref. 13, 14). TPO, a 933 amino-acid residues polypeptide, contains ectodomain, single membrane-spanning region and short intracellular tail (15). The dominant part of the protein, called ectodomain, consists of three distinct domains, namely MPO-like domain with high homology to myeloperoxidase (residues 142-738), CCP-like domain similar to complement control protein (residues 739-795), and EGF-like domain that is homologous to the epidermal growth factor (residues 796-841) (15). A three-dimensional computer model of TPO ectodomain was constructed in our laboratory, however, the localization of all TPO domains relatively to each other is still unknown (15). Till now the attempts to solve the TPO structure by X-ray crystallography ended in failure (16, 17). On the contrary, the crystal structure of other two members of the animal peroxidase family, namely MPO (18, 19) and LPO (20), has been already determined.
Post-translational modifications
The TPO molecule undergoes a few modifications while biosynthesis, that are glycosylation, heme incorporation, propeptide removal, and dimer formation (fig. 1). TPO ectodomain contains five predicted N-glycosylation sites in the amino-acid sequence (Asn129, Asn307, Asn342, Asn478, and Asn569) (6-8). However, glycosylation at position 478 probably does not occur because the X residue of the Asn-X-Thr consensus sequence is proline (21). It is well documented that N-linked oligosaccharides are added to the nascent protein in the endoplasmic reticulum (ER) (reviewed in Ref. 22). At this stage, the glycoprotein contains high mannose oligosaccharides that are sensitive to endoglycosidase H (Endo H) digestion. During the passage through the Golgi apparatus, glycoprotein undergoes a final process of N-glycosylation resulting in the transformation of Endo H-sensitive high mannose oligosaccharides to Endo H–resistant complex oligosaccharides. Thus, susceptibility to Endo H digestion is used to distinguish between high-mannose and complex oligosaccharides and help in differentiation whether the protein is before or after transit through the Golgi apparatus (reviewed in Ref. 22).
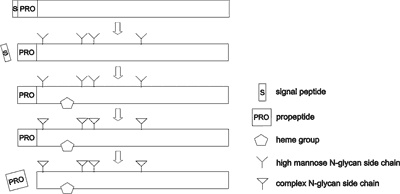
Fig. 1. TPO processing scheme. High mannose oligosaccharide side chains and heme group are incorporated to the TPO translation product in the endoplasmic reticulum. Signal peptide cleavage occurs also in this compartment (like in MPO) (reviewed in Ref. 47) or later during TPO transport to the cell surface (no data available). Some O-glycan side chains (not shown in this scheme), as well as complex N-glycan chains, are added to the TPO polypeptide in the Golgi apparatus. The propeptide is removed after its transit through the Golgi apparatus but before it reaches the plasma membrane. Mature TPO forms dimers (not shown in this scheme), however, the localization of dimerisation process in the cell is still unknown.
N-glycans content in human TPO was debated. The hypothesis, that TPO acquires complex oligosaccharides during maturation, was confirmed in research on TPO purified from human thyroid (23, 24), full-length TPO produced in insect (25) and CHO cells (26, 27), TPO ectodomain expressed in insect (24, 28) and CHO cells (28). Deglycosylation analysis of intracellular and cell surface fractions of TPO expressed in CHO cells showed that, inside the cell, this protein contains only high mannose-type structures, whereas TPO at plasma membrane bears complex-type structures (26). However, there are some reports suggesting that TPO contains only high mannose structures regardless of whether the protein is expressed in CHO cells (29) or is purified from thyroid gland (30). Additionally, other studies showed that TPO exiting ER does not necessarily acquire endoglycosidase H resistance (31). Human TPO produced in MDCK (Madin-Darby Canine Kidney) cells was Endo H-sensitive inside the cell and at the cell membrane. The same protein transfected to PC Cl3 (rat thyrocytes) was predominantly sensitive (about 75%) to Endo H digestion (being integrated to the plasma membrane) indicating the lack of conversion to complex carbohydrates upon passage through the Golgi complex (31). Nevertheless, in the same study it was observed that TPO ectodomain secreted to the medium by CHO cells was resistant to the endoglycosidase H digestion. The phenomenon of not acquiring Endo H resistance upon trafficking through the Golgi was earlier observed and it may be explained by the inaccessibility of high mannose oligosaccharides in the tertiary and quaternary structure (32). However, it needs explanation why in some studies the modification of these inaccessible high mannose N-glycans is possible.
N-linked oligosaccharide moieties serve various functions, e.g., stabilize the proteins against denaturation and proteolysis, improve solubility, modulate immune response, regulate protein turnover, stabilize the structure of proteins, and promote their adequate sorting and quality control (reviewed in Ref. 22, 33). The role of these units in TPO was examined. After Endo H treatment, the specific activity of TPO purified from human thyroid, measured by guaiacol and iodide oxidation assays, was reduced about 4 times in comparison to non-deglycosylated protein (30). The authors confirmed that the aggregated state of the molecule, that could be induced by the glycans removal, was not responsible for this decrease (30). They assigned this reduction to the modification of the molecule’s conformation at the enzymatic site (30). Kiso et al. (23) also demonstrated that enzymatic activity of native TPO treated with endoglycosidase H was significantly lower, when measured with guaiacol assay, but no effect was observed in the iodide oxidation assay. In further studies, the inhibition of N-glycosylation using tunicamycin (Tu) reduced by half enzymatic activity of human TPO at the cell surface in CHO cells (26). When the cells were treated with deoxymannojirimycin (dMM) which leads only to high mannose-type structures formation, the activity was not (or only slightly) reduced (26). The cell surface expression of newly synthesized TPO, after treatment with Tu, was almost totally blocked (by 95%) what may explain the decrease in enzymatic activity. All in all, it seems that N-glycans play an essential role in the intracellular trafficking and enzymatic activity of TPO. Additionally, the reactivity of unglycosylated TPO with anti-TPO antibodies, namely human autoimmune antibodies (23, 29, 30), mouse monoclonal antibodies (26, 30), and rabbit polyclonal antibodies (30), was determined. The conclusion was that the tertiary structure of epitopes recognized by anti-TPO antibodies is not markedly changed (23, 29, 30), however, the opposite observations were made by Fayadat et al. (26). Finally, as will be discussed later, N-linked oligosaccharides mediate the interaction between TPO and molecular chaperones (calnexin, calreticulin, and BiP) (34, 35).
The incorporation of O-glycans to human TPO was also analyzed. TPO purified from human thyroid (23), full-length human TPO expressed both in insect (25) and CHO cells (26) undergoes O-glycosylation. Opposite conclusions were drawn from the experiments conducted on full-length human TPO (29) and human TPO ectodomain (28), both expressed in CHO cells. In more detailed study, after separation of intracellular and cell-surface TPO, it was shown that only TPO localized in the latter fraction bears O-glycans (26). This finding was in agreement with the fact that O-glycosylation process occurs in the Golgi apparatus (reviewed in Ref. 33). O-glycans removal from native TPO, isolated from human thyroid gland, probably has no impact on enzymatic activity and interaction with autoantibodies (23). The treatment of the TPO-expressing CHO cells with phenyl-α-GalNAc, a specific inhibitor of O-glycosylation, only slightly altered TPO traffic to the cell surface and activity of this protein, nevertheless, the role of O-linked carbohydrates in the protection of TPO against proteolysis cannot be excluded (26).
The nature of the heme prosthetic group of TPO is still unclear. It was postulated that hog TPO contains other heme group than ferriprotoporphirin IX (36, 37). In other works, it was suggested that ferriprotoporphirin IX or other closely related porphyrin might be incorporated to the molecule of bovine (38) and hog TPO (39, 40). The probable localization in the TPO structure of proximal histidine (linked to the iron centre of the heme) and distal histidine (situated close to the peroxide binding pocket) was firstly proposed by Kimura and Ikeda-Saito (41). They indicated His407 or His414 as the candidates for the proximal and His494 or His586 as the probable distal ligand. More recently, His494 (42, 43) and His239 (43) were pointed out as possible proximal and distal histidine residues, respectively. It is hypothesized that Glu399 and Asn238 are bound covalently to the heme prosthetic group through ester linkage (43, reviewed in Ref. 44).
Powyżej zamieściliśmy fragment artykułu, do którego możesz uzyskać pełny dostęp.
Mam kod dostępu
- Aby uzyskać płatny dostęp do pełnej treści powyższego artykułu albo wszystkich artykułów (w zależności od wybranej opcji), należy wprowadzić kod.
- Wprowadzając kod, akceptują Państwo treść Regulaminu oraz potwierdzają zapoznanie się z nim.
- Aby kupić kod proszę skorzystać z jednej z poniższych opcji.
Opcja #1
24 zł
Wybieram
- dostęp do tego artykułu
- dostęp na 7 dni
uzyskany kod musi być wprowadzony na stronie artykułu, do którego został wykupiony
Opcja #2
59 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 30 dni
- najpopularniejsza opcja
Opcja #3
119 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 90 dni
- oszczędzasz 28 zł
Piśmiennictwo
1. Czarnocka B, Ruf J, Ferrand M et al.: Purification of the human thyroid peroxidase and its identification as the microsomal antigen involved in autoimmune thyroid diseases. FEBS Lett 1985; 190: 147-52.
2. Ruf J, Carayon P: Structural and functional aspects of thyroid peroxidase. Arch Biochem Biophys 2006; 445: 269-77.
3. McLachlan SM, Rapoport B: Thyroid peroxidase as an autoantigen. Thyroid 2007; 17: 939-48.
4. Hadj-Kacem H, Rebuffat S, Mnif-Féki M et al.: Autoimmune thyroid diseases: genetic susceptibility of thyroid-specific genes and thyroid autoantigens contributions. Int J Immunogenet 2009; 36: 85-96.
5. Czarnocka B: Thyroperoxidase, thyroglobulin, Na(+)/I(-) symporter, pendrin in thyroid autoimmunity. Front Biosci 2011; 16: 783-802.
6. Kimura S, Kotani T, McBride OW et al.: Human thyroid peroxidase: complete cDNA and protein sequence, chromosome mapping, and identification of two alternately spliced mRNAs. Proc Natl Acad Sci U S A 1987; 84: 5555-9.
7. Libert F, Ruel J, Ludgate M et al.: Complete nucleotide sequence of the human thyroperoxidase-microsomal antigen cDNA. Nucleic Acids Res 1987; 15: 6735.
8. Magnusson RP, Chazenbalk GD, Gestautas J et al.: Molecular cloning of the complementary deoxyribonucleic acid for human thyroid peroxidase. Mol Endocrinol 1987; 1: 856-61.
9. de Vijlder JJ, Dinsart C, Libert F: Regional localization of the gene for thyroid peroxidase to human chromosome 2pter-p12. Cytogenet Cell Genet 1988; 47: 170-2.
10. McLachlan SM, Rapoport B: The molecular biology of thyroid peroxidase: cloning, expression and role as autoantigen in autoimmune thyroid disease. Endocr Rev 1992; 13: 192-206.
11. Ohtaki S, Nakagawa H, Nakamura M et al.: Thyroid peroxidase: experimental and clinical integration. Endocr J 1996; 43: 1-14.
12. Xing Y, Kuang A: Development of studies of TPO gene and its application in nuclear medicine. Nucl Med Commun 2003; 24: 853-6.
13. O’Brien PJ: Peroxidases. Chem Biol Interact 2000; 129: 113-39.
14. Furtmüller PG, Zederbauer M, Jantschko W et al.: Active site structure and catalytic mechanisms of human peroxidases. Arch Biochem Biophys 2006; 445: 199-213.
15. Hobby P, Gardas A, Radomski R et al.: Identification of an immunodominant region recognized by human autoantibodies in a three-dimensional model of thyroid peroxidase. Endocrinology 2000; 141: 2018-26.
16. Gardas A, Sohi MK, Sutton BJ.: Purification and crystallisation of the autoantigen thyroid peroxidase from human Graves’ thyroid tissue. Biochem Biophys Res Commun 1997; 234: 366-70.
17. Hendry E, Taylor G, Ziemnicka K et al.: Recombinant human thyroid peroxidase expressed in insect cells is soluble at high concentrations and forms diffracting crystals. J Endocrinol 1999; 160: R13-5.
18. Zeng J, Fenna RE: X-ray crystal structure of canine myeloperoxidase at 3 A resolution. J Mol Biol 1992; 226: 185-207.
19. Fiedler TJ, Davey CA, Fenna RE: X-ray crystal structure and characterization of halide-binding sites of human myeloperoxidase at 1.8 A resolution. J Biol Chem 2000; 275: 11964-71.
20. Singh AK, Singh N, Sharma S et al.: Crystal structure of lactoperoxidase at 2.4 A resolution. J Mol Biol 2008; 376: 1060-75.
21. Roitsch T, Lehle L: Structural requirements for protein N-glycosylation. Influence of acceptor peptides on cotranslational glycosylation of yeast invertase and site-directed mutagenesis around a sequon sequence. Eur J Biochem 1989; 181: 525-9.
22. Helenius A, Aebi M: Roles of N-linked glycans in the endoplasmic reticulum. Annu Rev Biochem 2004; 73: 1019-49.
23. Kiso Y, Furmaniak J, Morteo C et al.: Analysis of carbohydrate residues on human thyroid peroxidase (TPO) and thyroglobulin (Tg) and effects of deglycosylation, reduction and unfolding on autoantibody binding. Autoimmunity 1992; 12: 259-69.
24. Grennan Jones F, Wolstenholme A, Fowler S et al.: High-level expression of recombinant immunoreactive thyroid peroxidase in the High Five insect cell line. J Mol Endocrinol 1996; 17: 165-74.
25. Fan JL, Patibandla SA, Kimura S et al.: Purification and characterization of a recombinant human thyroid peroxidase expressed in insect cells. J Autoimmun 1996; 9: 529-36.
26. Fayadat L, Niccoli-Sire P, Lanet J et al.: Human thyroperoxidase is largely retained and rapidly degraded in the endoplasmic reticulum. Its N-glycans are required for folding and intracellular trafficking. Endocrinology 1998; 139: 4277-85.
27. Le Fourn V, Ferrand M, Franc JL: Endoproteolytic cleavage of human thyroperoxidase: role of the propeptide in the protein folding process. J Biol Chem 2005; 280: 4568-77.
28. Guo J, McLachlan SM, Hutchison S et al.: The greater glycan content of recombinant human thyroid peroxidase of mammalian than of insect cell origin facilitates purification to homogeneity of enzymatically protein remaining soluble at high concentration. Endocrinology 1998; 139: 999-1005.
29. Foti D, Rapoport B: Carbohydrate moieties in recombinant human thyroid peroxidase: role in recognition by antithyroid peroxidase antibodies in Hashimoto’s thyroiditis. Endocrinology 1990; 126: 2983-8.
30. Giraud A, Franc JL, Long Y et al.: Effects of deglycosylation of human thyroperoxidase on its enzymatic activity and immunoreactivity. J Endocrinol 1992; 132: 317-23.
31. Zhang X, Arvan P: Cell type-dependent differences in thyroid peroxidase cell surface expression. J Biol Chem 2000; 275: 31946-53.
32. Cals MM, Guenzi S, Carelli S et al.: IgM polymerization inhibits the Golgi-mediated processing of the mu-chain carboxy-terminal glycans. Mol Immunol 1996; 33: 15-24.
33. Ungar D: Golgi linked protein glycosylation and associated diseases. Semin Cell Dev Biol 2009; 20: 762-9.
34. Fayadat L, Siffroi-Fernandez S, Lanet J et al.: Calnexin and calreticulin binding to human thyroperoxidase is required for its first folding step(s) but is not sufficient to promote efficient cell surface expression. Endocrinology 2000; 141: 959-66.
35. Le Fourn V, Siffroi-Fernandez S, Ferrand M e al.: Competition between calnexin and BiP in the endoplasmic reticulum can lead to the folding or degradation of human thyroperoxidase. Biochemistry 2006; 45: 7380-8.
36. Taurog A, Lothrop ML, Estabrook RW: Improvements in the isolation procedure for thyroid peroxidase: nature of the heme prosthetic group. Arch Biochem Biophys 1970; 139: 221-9.
37. Rawitch AB, Taurog A, Chernoff SB et al.: Hog thyroid peroxidase: physical, chemical, and catalytic properties of the highly purified enzyme. Arch Biochem Biophys 1979; 194: 244-57.
38. Krinsky MM, Alexander NM: Thyroid peroxidase. Nature of the heme binding to apoperoxidase. J Biol Chem 1971; 246: 4755-8.
39. Hosoya T, Morrison M: A study of the hemoproteins of thyroid microsomes with emphasis on the thyroid peroxidase. Biochemistry 1967; 6: 1021-6.
40. Ohtaki S, Nakagawa H, Nakamura S et al.: Characterization of hog thyroid peroxidase. J Biol Chem 1985; 260: 441-8.
41. Kimura S, Ikeda-Saito M: Human myeloperoxidase and thyroid peroxidase, two enzymes with separate and distinct physiological functions, are evolutionarily related members of the same gene family. Proteins 1988; 3: 113-20.
42. Bikker H, Baas F, De Vijlder JJ: Molecular analysis of mutated thyroid peroxidase detected in patients with total iodide organification defects. J Clin Endocrinol Metab 1997; 82: 649-53.
43. Taurog A, Wall M: Proximal and distal histidines in thyroid peroxidase: relation to the alternatively spliced form, TPO-2. Thyroid 1998; 8: 185-91.
44. Taurog A: Molecular evolution of thyroid peroxidase. Biochimie 1999; 81: 557-62.
45. Fayadat L, Niccoli-Sire P, Lanet J: Role of heme in intracellular trafficking of thyroperoxidase and involvement of H2O2 generated at the apical surface of thyroid cells in autocatalytic covalent heme binding. J Biol Chem 1999; 274: 10533-8.
46. Chen YJ, Inouye M: The intramolecular chaperone-mediated protein folding. Curr Opin Struct Biol 2008; 18: 765-70.
47. Hansson M, Olsson I, Nauseef WM: Biosynthesis, processing, and sorting of human myeloperoxidase. Arch Biochem Biophys 2006; 445: 214-24.
48. Hamada N, Grimm C, Mori H et al.: Identification of a thyroid microsomal antigen by Western blot and immunoprecipitation. J Clin Endocrinol Metab 1985; 61: 120-8.
49. Hamada N, Portmann L, DeGroot LJ: Characterization and isolation of thyroid microsomal antigen. J Clin Invest 1987; 79: 819-25.
50. Hamada N, Jaeduck N, Portmann L et al.: Antibodies against denatured and reduced thyroid microsomal antigen in autoimmune thyroid disease. J Clin Endocrinol Metab 1987; 64: 230-8.
51. Baker JR, Arscott P, Johnson J: An analysis of the structure and antigenicity of different forms of human thyroid peroxidase. Thyroid 1994; 4: 173-8.
52. Kaufman KD, Rapoport B, Seto P et al.: Generation of recombinant, enzymatically active human thyroid peroxidase and its recognition by antibodies in the sera of patients with Hashimoto’s thyroiditis. J Clin Invest 1989; 84: 394-403.
53. Nishikawa T, Rapoport B, McLachlan SM: Exclusion of two major areas on thyroid peroxidase from the immunodominant region containing the conformational epitopes recognized by human autoantibodies. J Clin Endocrinol Metab 1994; 79: 1648-54.
54. McDonald DO, Pearce SH: Thyroid peroxidase forms thionamide-sensitive homodimers: relevance for immunomodulation of thyroid autoimmunity. J Mol Med 2009; 87: 971-80.
55. Kuliawat R, Lisanti MP, Arvan P: Polarized distribution and delivery of plasma membrane proteins in thyroid follicular epithelial cells. J Biol Chem 1995; 270: 2478-82.
56. Penel C, Gruffat D, Alquier C et al.: Thyrotropin chronically regulates the pool of thyroperoxidase and its intracellular distribution: a quantitative confocal microscopic study. J Cell Physiol 1998; 174: 160-9.
57. Kuliawat R, Ramos-Casta?eda J, Liu Y et al.: Intracellular trafficking of thyroid peroxidase to the cell surface. J Biol Chem 2005; 280: 27713-8.
58. Giraud A, Lejeune PJ, Barbaria J et al.: A plasminogen-like protease in thyroid rough microsomes degrades thyroperoxidase and thyroglobulin. Endocrinology 2007; 148: 2886-93.
59. Giraud A, Dicristofaro J, De Micco C et al.: A plasminogen-like protein, present in the apical extracellular environment of thyroid epithelial cells, degrades thyroglobulin in vitro. Biochem Biophys Res Commun 2005; 338: 1000-4.
60. Coe H, Michalak M: ERp57, a multifunctional endoplasmic reticulum resident oxidoreductase. Int J Biochem Cell Biol 2010; 42: 796-9.
61. Wang D, De Deken X, Milenkovic M et al.: Identification of a novel partner of duox: EFP1, a thioredoxin-related protein. J Biol Chem 2005; 280: 3096-103.
62. Scott DA, Wang R, Kreman TM et al.: The Pendred syndrome gene encodes a chloride-iodide transport protein. Nat Genet 1999; 21: 440-3.
63. Riesco-Eizaguirre G, Santisteban P: A perspective view of sodium iodide symporter research and its clinical implications. Eur J Endocrinol 2006; 155: 495-512.
64. Song Y, Driessens N, Costa M et al.: Roles of hydrogen peroxide in thyroid physiology and disease. J Clin Endocrinol Metab 2007; 92: 3764-73.
65. Ohye H, Sugawara M: Dual oxidase, hydrogen peroxide and thyroid diseases. Exp Biol Med (Maywood) 2010; 235: 424-33.
66. Song Y, Ruf J, Lothaire P et al.: Association of duoxes with thyroid peroxidase and its regulation in thyrocytes. J Clin Endocrinol Metab 2010; 95: 75-82.
67. Ameziane-El-Hassani R, Morand S, Boucher JL et al.: Dual oxidase-2 has an intrinsic Ca2+-dependent H2O2 – generating activity. J Biol Chem 2005; 280: 30046-54.
68. Fortunato RS, Lima de Souza EC, Ameziane-el Hassani R et al.: Functional consequences of dual oxidase-thyroperoxidase interaction at the plasma membrane. J Clin Endocrinol Metab 2010; 95: 5403-11.
69. Ehrenshaft M, Mason RP: Protein radical formation on thyroid peroxidase during turnover as detected by immuno-spin trapping. Free Radic Biol Med 2006; 41: 422-30.