© Borgis - Medycyna Rodzinna 4/2004, s. 187-192
Iwona Urbanowicz, Jolanta Stacherzak-Pawlik, Mieczysław Woźniak
Aktywność telomerazy i długość telomerów w rozrostach układu chłonnego
Telomerase activity and telomere length in lymphoproliferative disorders
z Katedry i Zakładu Analityki Medycznej AM we Wrocławiu
Kierownik Katedry: prof. dr hab. m. Mieczysław Woźniak
Summary
Telomerase is a telomere – synthesing reverse transcriptase. that can compensate for the loss of telomere associated with cell divisions. Telomerase activity has been found in the majority of cancer, but absent in the most normal cells .However telomerase activity is detectable at low levels in normal human T and B cells, increases in hematopoietic progenitor cells upon their proliferation and differentiation. Shortened telomeres and high telomerase activity almost always correlates with disease severity in lymphoproliferative disorders. Measurement of telomere length and telomerase activity might be useful to monitor disease condition. and as well prognostic marker in lymphoproliferative disorders.

Do podstawowych przyczyn transformacji nowotworowej należą zaburzenia ekspresji genów, których produkty białkowe biorą udział w regulacji cyklu komórkowego i przenoszeniu sygnału wewnątrz komórki. Mutacje tych genów powodują inaktywacje białek, co pozbawia komórki niezbędnych inhibitorów proliferacji. W poszukiwaniu mechanizmów regulujących aktywność podziałową komórek zwrócono w ostatnich latach uwagę na zagadnienia dotyczące replikacji chromosomów. Odkrycie na końcach eukariotycznych chromosomów powtarzalnych sześcionukleotydowych sekwencji określanych mianem telomerów oraz regulującej ich długość telomerazy czyni z nich interesujący obiekt badań.
Określenie roli telomerów i telomerazy w podstawowych procesach życiowych komórki było możliwe dzięki poznaniu ich struktury.
Telomery są dynamicznymi, wyspecjalizowanymi strukturami nukleoproteinowymi występującymi na końcach eukariotycznych chromosomów. Składają się z powtarzających się sekwencji niekodującego DNA bogatego w zasady guaninowe – G (nić 3´) i odpowiednio w cytozynowe – C (nić 5´) z jednoniciowym końcem 3´ bogatym w G. Telomery tworzą wielkie końcowe pętle widoczne pod mikroskopem elektronowym, nazywane pętlami t (od „telomerów”, t – loop) (1, 2). Dwuniciowy DNA tworzy pętlę t, natomiast jednoniciowe końce o długości od 50 do ok. 150 nukleotydów układają się wewnątrz dwuniciowego DNA, tworząc małe pętle D (displacement, d-loop) (3). Powstanie pętli t tłumaczy możliwość rozróżniania końców chromosomów od uszkodzeń DNA. Ich struktura i ich długość są w nieznacznym stopniu zróżnicowane gatunkowo.
Ludzkie telomery mają długość do 25 tys. bp (base pairs, par zasad) (4) i są zbudowane z powtarzających się heksanukleotydów TTAGGG. Liczba powtórzeń różni się pomiędzy chromosomami tej samej komórki, w obrębie populacji pozostaje jednak w ściśle określonym przedziale (5).
Telomery zabezpieczają DNA przed rearanżacjami prowadzącymi do niestabilności genomu i zmian w kariotypie. Chronią one chromosomy przed zmianą struktury i ich fuzją z innymi chromosomami, biorą udział w przestrzennej organizacji jądra komórkowego, segregacji chromosomów podczas podziału komórki i w regulacji transkrypcji genów zlokalizowanych w ich pobliżu (5). Ponadto podkreśla się ich rolę jako magazynów białek uczestniczących w naprawie DNA. Chroniąc końce chromosomów przed działaniem nukleaz nie dopuszczają do procesu apoptozy.
W przedstawione powyżej procesy zaangażowane są białka związane z jedno- lub dwuniciowym telomerowym DNA. Jedynymi znanymi komponentami białkowymi telomerów u ssaków są białka: TRF1 i TRF2 – czynniki wiążące się do podwójnych powtórzeń TTAGGG (duplex TTAGGG repeat – binding factors, TRF) w obrębie pętli t. Białko TRF1 jest negatywnym regulatorem długości telomerów i zapobiega wiązaniu się telomerazy (3). Białko TRF2 wpływa też na jednoniciowe końce, spinając je i zapobiegając fuzji końców. Jego brak powoduje utratę końców G, indukcję drogi zależnej od białka p53 i fuzję końców chromosomów (6, 7). W okolicy porów jądrowych i centrosomu odkryto obecność tankyrazy (TRF1-interacting, ANKYrin-related ADP-ribose polymerase) (8), która jest inhibitorem białka TRF1 i może stanowić jeden z elementów regulacyjnych procesu tworzenia i utrzymania pętli t (9). Skracanie telomerów oraz utrata funkcji w komórkach pozbawionych telomerazy może wynikać z rozbicia struktury t (1, 2).
Telomeraza jest enzymem o charakterze odwrotnej transkryptazy, która poprzez zdolność syntetyzowania telomerów zabezpiecza końce chromosomów przed ich nadmiernym skracaniem się w trakcie podziałów komórkowych oraz stabilizuje ich strukturę. Telomeraza ludzka jest kompleksem zbudowanym z:
– białkowej podjednostki katalitycznej, polimerazy DNA zależnej od RNA nazywanej telomerazową odwrotną transkryptazą – hTERT (human telomerase reverse transcriptase),
– matrycy RNA (hTR, human template RNA),
– białek zasocjowanych z telomerazą: TRF1 i TRF2 (TTAGGG repeat binding factors) opisanych powyżej.
W prawidłowych komórkach organizmu ludzkiego aktywność telomerazy jest wykrywana w czasie ich rozwoju embrionalnego i stopniowo obniża się w trakcie różnicowania tkankowego. Wysoka aktywność telomerazy jest zachowana w komórkach rozrodczych, aktywowanych limfocytach T i B oraz komórkach macierzystych szpiku kostnego (3, 10). Prawidłowe komórki pozbawione aktywności telomerazy nie wykazują również ekspresji białkowej podjednostki katalitycznej hTERT, pomimo iż, wykrywa się w nich ekspresję hTR lub białek związanych z telomerazą. Wydaje się, więc, że czynnikiem odpowiedzialnym za aktywność telomerazy jest podjednostka katalityczna hTERT i regulacja jej ekspresji jest kluczowa dla wydłużenia potencjału proliferacyjnego komórki. Regulacja ekspresji hTERT może przebiegać na poziomie posttranskrypcyjnym i posttranslacyjnym (11, 12, 13). Wzrost ekspresji hTERT na poziomie transkrypcyjnym zależy m.in. od bezpośredniej interakcji produktu onkogenu c-myc, hormonów steroidowych tj. estrogenu, progesteronu z regionem promotorowym dla genu hTERT. Do czynników, które obniżają ekspresję hTERT należą: czynniki transkrypcyjne (E2F-1, Mad), białko supresorowe – p53, inhibitor kinaz cyklinozależnych p16INK4a. W regulacji posttranslacyjnej zasadniczą rolę odgrywają kinazy białkowe serynowo-treoninowe. Do prawidłowej aktywności enzymu niezbędna jest fosforylacja białkowej podjednostki hTERT (3).
BADANIE DŁUGOŚCI TELOMERÓW
Podstawową metodą oceny długości telomerów był dotąd Southern blot, w którym otrzymuje się fragmenty TRF (telomere restriction fragment) przy użyciu enzymów restrykcyjnych. dla rejonów subtelomerycznych, np. Rsa I, Msp I. TRF są rozpoznawane za pomocą sond molekularnych znakowanych izotopowo np. fosforem 32P. Ze względu na jej czasochłonność została wyparta przez metodę cytometrii przepływowej wykonywaną w ciągu kilku godzin. W ocenie cytometrycznej telomerów istotnym elementem jest hybrydyzacja in situ sondą PNA (peptide nucleic acid) sprzężoną z fluorochromem FITC. Możliwe jest zbadanie tysięcy komórek w ciągu kilkudziesięciu sekund oraz ocena długości telomerów w obrębie określonych subpopulacji komórek w połączeniu z oceną cyklu komórkowego. Wyniki badań telomerów wykazują wysoką korelację z pomiarami fragmentów TRF (14).
Oznaczenie telomerów w cytometrze przepływowym wymaga określonej procedury: w pierwszej kolejności ma miejsce wstępna selekcja pozytywna lub negatywna badanych komórek (separacja magnetyczna, rozetki), następnie denaturacja DNA i hybrydyzacja. Do komórek zawieszonych w roztworze hybrydyzacyjnym, dodaje się sondę PNA telomerowego (CCCTAA)3 sprzężoną z FITC. W celu oceny fluorescencji tła ocenia się negatywną kontrolę, którą są komórki zawieszone w mieszaninie hybrydyzacyjnej bez PNA. Po wypłukaniu komórek w roztworze post-hybrydyzacyjnym barwi się jądra jodkiem propydyny łącznie z RNAzą. Jeśli badane komórki charakteryzują się znaczną proliferacją, konieczne jest skorygowanie cyklu wyników przez wyłączenie komórek będących w fazie S oraz G2/M cyklu komórkowego.
Badanie telomerazy polega na ocenie jej aktywności enzymatycznej, jak również na badaniu poziomu ekspresji elementów składowych kompleksu telomerazy (hTR, hTERT) (14).
Aktywność telomerazy mierzona jest zdolnością lizatu komórkowego do syntetyzowania in vitro sekwencji telomerów. Telomeraza z lizatu komórkowego wydłuża oligonukleotydowe primery TS – 5´-AATCCGTCGAGCAGAGTT-3´ syntetyzując powtarzalne sześcionukleotydowe sekwencje. Metoda ta jest wystarczająca dla oceny aktywności enzymu w hodowlach komórkowych, ale nie jest dostatecznie czuła dla małej ilości materiału klinicznego. Dlatego aktywność telomerazy ocenia się obecnie za pomocą zestawu TRAP (telomeric repeat amplification protocol) (ryc. 1) (14).
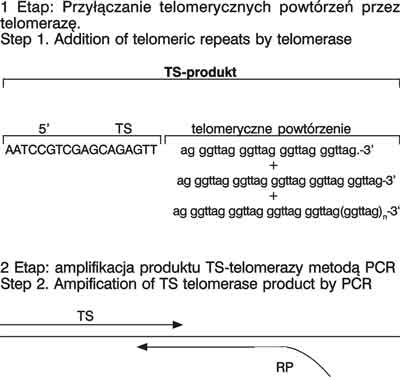
Ryc. 1. Schemat oznaczania telomerazy metodą TRAP (TS –sonda 5´<font face="Symbol">(r)3´),
RP (reverse primer) 3´<font face="Symbol">(r)5´),
TRAPeze telomerase Detection Kit Assay Scheme (TS – primer.5´<font face="Symbol">(r)3´),
RP(reverse primer) 3´<font face="Symbol">(r)5´).
Badanie to charakteryzuje się wysoką czułością, a wymagana liczba komórek mieści się od 100 do 1000 komórek w badanym materiale. Metoda TRAP oparta jest na syntezie telomeru – przyłączenia heksamerów GGTTAG do 3´ końca substratu oligonukleotydowego (n) razy i późniejszej amplifikacji pierwotnego produktu w reakcji PCR (reakcja łańcuchowej polimerazy). Zastosowanie PCR pozwala na wykrycie śladowych ilości produktu, a tym samym potwierdza aktywność enzymu. Produkty PCR są następnie poddawane rozdziałwi elektroforetycznemu w żelu poliakrylamidowym, gdzie przedstawiają się jako drabinka o wielkości od 50 + 6n bp. Otrzymane prążki uwidacznia się drogą autoradiografii po uprzednim znakowaniu oligonukleotydowych primerów izotopem np. fosforem 32P, bądź poprzez barwienie żelu np. bromkiem etydyny. Gęstość prążków proporcjonalną do aktywności enzymatycznej telomerazy ocenia się densytometrycznie (15).
BADANIE EKSPRESJI PODJEDNOSTKI KATALITYCZNEJ TELOMERAZY HTERT
Czynnikiem odpowiedzialnym za aktywność telomerazy jest podjednostka katalityczna hTERT. Jej ekspresję możemy zbadać:
a) za pomocą metody RT-PCR (reakcja odwrotnej transkrypcji – RT i reakcji PCR). Metoda ta polega na syntezie w warunkach in vitro komplementarnego DNA na matrycy wyizolowanego wcześniej z badanych komórek mRNA specyficznego dla hTERT, a następnie reakcji amplifikacji za pomocą PCR z użyciem primerów hTERT i z koamplifikacją GAPDH lub aktyny jako kontroli wewnętrznej. Produkty reakcji PCR mogą być wizualizowane w żelu agarozowym zawierającym bromek etydyny i oceniane densytometrycznie;
b) w cytometrze przepływowym w połączeniu z oceną ekspresji innych antygenów komórkowych. Zastosowanie tego protokołu pozwala wyróżnić w całym układzie badanych komórek subpopulacje o różnej, często niskiej ekspresji enzymu, których wartość zostaje uśredniona w innych metodach. Wartość ekspresji hTERT otrzymujemy po odjęciu od średniej fluorescencji badanej populacji komórek fluorescencję kontroli (14).
Telomeraza jest obecna w większości komórek nowotworowych człowieka. Komórki nowotworowe uaktywniają telomerazę, a jej pojawienie się jest najprawdopodobniej niezbędnym etapem dla ich przeżycia. Wydaje się, że nawet obecność mutacji w obrębie genów kodujących kilka protoonkogenów (ras, myc) oraz genów supresorowych (p53, RB-1) nie wystarcza do immortalizacji. Przy zbyt krótkich telomerach dochodzi do stopniowej kumulacji coraz większej liczby defektów genetycznych (fuzji chromosomów, translokacji) uniemożliwiających przetrwanie nawet „najbardziej stransformowanym” komórkom nowotworowym (3).
Z każdym podziałem komórki dochodzi do stopniowego skracania się telomerów. Proces ten bywa porównywany do zegara mitotycznego odmierzającego czas życia komórek. Zbyt krótkie telomery są sygnałem informującym o starzeniu się i są rozpoznawane przez mechanizmy regulujące cykl komórkowy jako moment sygnalizujący konieczność wstrzymania dalszych podziałów. Wiele komórek ma aktywność telomerazową w stadium embrionalnym i traci ją podczas różnicowania tkankowego. Istnieją jednak komórki, które zachowują aktywność enzymu do końca życia osobniczego. Są to komórki rozrodcze, komórki macierzyste oraz limfocyty podczas ich podziałów, różnicowania i aktywacji.
Interesujący model do badania proliferacji komórkowej tak ze względów poznawczych jak i klinicznych stanowi układ chłonny. Populacje komórek nowotworowych rozrostów limforetikularnych odpowiadają morfologicznie i immunologicznie różnym stadiom ontogenezy układu chłonnego oraz wykazują duże zróżnicowanie w zakresie aktywności podziałowej, nasilenia apoptozy oraz zaawansowania procesu dojrzewania.
Nieprzerwana proliferacja komórek hematopoetycznych jest wymagana do utrzymania hematopoezy przez całe życie. Krwiotwórcze komórki macierzyste (hematopoietic stem cells) mogą utrzymywać lub nawet wydłużają telomery poprzez aktywność telomerazy. W przybliżeniu 70% krwiotwórczych komórek macierzystych szpiku kostnego wykazuje wykrywalny poziom aktywności telomerazy (16). Wyniki badań Morrisona i wsp. (1996) wskazują, iż poziom tej aktywności zależy od stadium różnicowania komórek hematopoetycznych i w każdej populacji jest proporcjonalny do wielkości frakcji komórek mających zamiar się odnowić (17). Podobne wyniki otrzymał Chui i wsp. (1996) wykazując wysoką aktywność telomerazy we wczesnych ukierunkowanych komórkach szpiku kostnego (CD34+CD71+), natomiast niską w komórkach hematopoetycznych niezróżnicowanych o fenotypie (CD34+CD71loCD45RAlo) (18). Tak więc, w komórkach szpiku kostnego aktywność telomerazy jest wykrywana w komórkach już zaangażowanych w proces różnicowana, a jej brak lub niski poziom jest stwierdzany w komórkach najbardziej prymitywnych.
Zachowanie telomerów zależy od aktywności telomerazy. Analiza długości telomerów wykazała, że są one krótsze w komórkach macierzystych u osób dorosłych, niż w embrionalnej tkance wątroby czy we krwi pępowinowej. Również komórki macierzyste badane w szpiku kostnym dawców młodych i starszych wykazują różnice w długości telomerów. Są one dłuższe u osób młodszych (19).
Z pierwotnej macierzystej komórki hematopoetycznej wywodzą się m.in. limfocyty T i B. Podobnie jak w innych komórkach również w limfocytach T i B wraz z wiekiem obserwuje się stopniowe skracanie się telomerów. Limfocyty B tracą telomery w wolniejszym tempie w stosunku do limfocytów T: ok. 19 bp/rok dla B, a od 30 do 50 bp/rok w komórkach limfatycznych T (20). Zmiany w długości telomerów komórek limfatycznych występują już podczas rozwoju embrionalnego. Szybkie i duże skrócenie telomerycznego fragmentu DNA występuje między 27 a 32 tygodniem ciąży a następnie między 33 a 42 tygodniem jej trwania. Intensywnie postępująca utrata telomerów w limfocytach obserwowana jest w pierwszym roku życia osobniczego w porównaniu z następnymi latami (21).
Wysoka aktywność telomerazy jest stwierdzana na każdym etapie różnicowania się limfocytów T, natomiast jest słabo wykrywalna w dojrzałych limfocytach T krwi obwodowej, które są w większości w fazie spoczynkowej (G0). Wysoki poziom aktywności telomerazy był obserwowany w tymocytach, a niski w dojrzałych limfocytach CD4+ i CD8+ (22). Wyraźna modulacja aktywności telomerazy występuje w limfocytach poddanych aktywacji antygenami. Aktywacja limfocytów T krwi obwodowej w warunkach in vitro za pomocą fitohemaglutyniny (PHA) poprzez ich specyficzny receptor (TCR) lub za pomocą przeciwciał monoklonalnych anty-CD3, anty-CD3/CD28 prowadzi do bardzo dużego wzrostu aktywności enzymu w pierwszych dniach hodowli, a następnie obserwuje się stopniowy jej spadek, pomimo że komórki pozostają nadal w fazie proliferacji. Obserwowana aktywność telomerazy wykazuje ścisły związek z wysoką ekspresją podjednostki katalitycznej hTERT (22).
Powyżej zamieściliśmy fragment artykułu, do którego możesz uzyskać pełny dostęp.
Mam kod dostępu
- Aby uzyskać płatny dostęp do pełnej treści powyższego artykułu albo wszystkich artykułów (w zależności od wybranej opcji), należy wprowadzić kod.
- Wprowadzając kod, akceptują Państwo treść Regulaminu oraz potwierdzają zapoznanie się z nim.
- Aby kupić kod proszę skorzystać z jednej z poniższych opcji.
Opcja #1
24 zł
Wybieram
- dostęp do tego artykułu
- dostęp na 7 dni
uzyskany kod musi być wprowadzony na stronie artykułu, do którego został wykupiony
Opcja #2
59 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 30 dni
- najpopularniejsza opcja
Opcja #3
119 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 90 dni
- oszczędzasz 28 zł
Piśmiennictwo
1. Greider C.W.: Telomeres do D-loop - T-loop. Cell 1999, 97, 419-422. 2.Griffith J.D., et al.: Mammalian Telomeres End in a Large Duplex Loop. Cell 1999, 97, 503-514. 3.Nowis D., Gołąb J.: Rola telomerów i telomerazy w progresji nowotworów. Perspektywy diagnostyczne i terapeutyczne. Postępy Biologii Komórki 2001, 28, 243-261. 4.Greider C.W.: Telomere length regulation. Annu Rev. Biochem., 1996, 5, 337-365. 5.Urquidi V., et al.: Role of telomerase.in cell senescense and oncogenesis. Annu Rev. Med., 2000, 51, 65-79. 6.Karlseder J., et al.: p53 and ATM-dependent apoptosis induced by telomeres lacking TRF2. Science 1999, 283, 1321-1325. 7.van Steensel B., et al.: TRF2 protects human telomeres from end-to-end fusion´s. Cell 1998, 92, 401-413. 8.Dhaene K., et al.: Telomeres telomerase and cancer: an up-date. Virchows Arch., 2000, 437, 1-16. 9.d´Adda di Fanga F., et al.: Functions of poly (ADP-ribose) polymerase controlling telomere length and chromosomal stability. Nature Genet., 1999, 23, 76-80. 10.Liu K., et al.: Constitutive and regulated expression of telomerase reverse transcriptase (hTERT) in human lymphocytes. Proc. Natl. Acad. Sci. USA 1999, 96, 5147-5152. 11.Mauro L.J., Foster D.N.: Regulators of telomerase activity. Am. J. Respir. Cell. Mol. Biol., 2002, 26, 521-524. 12.Meyerson M., et al.: hEST2 the putative human telomerase catalytic subunit gene, is up-regulated in tumor cells and during immortalization. Cell., 1997, 97, 609-620. 13.Nilsson P., Mehle C., Remesk.: Telomerase activity in vivo in human malignant hematopoietic cells. Oncogene 1994, 9, 3043-8. 14.Dragan M., Roliński J.: Ocena długości telomerów i aktywności telomerazy za pomocą cytometrii przepływowej. Post. Biol. Kom., 2001, 28, 4 (471-481). 15.Nowak J., et al.: Rola telomerazy w rozwoju procesu nowotworowego. Postępy Biologii Komórki 2000, 27 nr 14 (143-150). 16.Weng N.P.: Interplay between telomere length and telomerase in human leukocyte differentiation and aging. J. Leukoc Biol., 2001, 70, 861-867. 17.Morrison S.J., et al.: Telomerase activity in hematopoietic cells is associated with self-renewal potential. Immunity 1996, 5, 207-216. 18.Chui C., et al.: Differential expression of telomerase activity in hematopoietic progenitors from adult human bone marrow. Stem. Cells., 1996, 14, 239-248. 19.Vaziri H., et al.: Evidence for a mitotic clock in human hematopoietic stem cells: loss of telomeric DNA with age. Proc. Natl. Acad. Sci. USA 1994, 91, 9857-9860. 20.Son N.H., et al.: Lineage-specific telomere shortening and unaltered capacity for telomerase expression in human T and B lymphocytes with age. J. Immunol., 2000, 165, 1191-1196. 21.Fridrich U., et al.: Telomeres in neonates: new insights in fetal hematopoiesis. Pediatr. Res., 2001, 49, 252-256. 22.Boussin F., et al.: Télomčres et télomérase dans les cellules hématopoďétiques et les hémopathies malignes. La Lettre du Canceroloque 1999, 8, 58. 23.Monteiro J., et al.: Shortened telomeres in clonally expanded CD28- CD8+ T cells imply a replicative history that is distinct from their CD28+CD8+ counterparts. J. Immunol., 1996, 156, 3587-3590. 24.Engelhardt M., et al.: Telomerase activity and telomere length in pediatric patients with malignancies undergoing chemotherapy. Leukemia 1998, 12, 13-24. 25.Uchida N., et al.: Correlation of telomerase activity with development and progression of adult T-cells leukemia. Leuk. Res., 1999, 23(3), 311-316. 26.Bechter O.E., et al.: Telomere length and telomerase activity predict survival in patients with B cell chronic lymphocytic leukemia. Cancer Research 1998, 58, 4918-4922. 27.Counter M., et al.: Telomerase activity in normal leukocytes and in hematologic malignancies. Blood 1995, 85, 2315-20. 28.Holt S.E., et al.: Regulation of telomerase activity in immortal cell lines. Molecular and Cellular Biology 1996, 16, 2932-2939. 29.Ballon G., et al.: Telomerase activity and clinical progression in chronic lymphoproliferative disorders of B-CLL lineage. Leukemia and Lymphoma 2001, 41, 35-45. 30.Trentin L., et al.: Telomerase activity in chronic lymphoproliferative disorders of B-cell lineage. Br. J. Hematol., 1999, 106, 662-668. 31.Brousset P., et al.: Telomerase activity in reactive and neoplastic lymphoid tissues: infrequent detection of activity in Hodgkin´s disease. Blood 1997, 89, 26-31. 32.Brousset P., et al.: Telomerase activity in Hodgkin´s disease. Leuk. Lymph., 1998, 30, 189-192. 33.Norrback K.F., et al.: Telomerase activity in Hodgkin´s disease. Blood 1998, 92, 587-73. 34.Chin L., et al.: p53 deficiency rescues the adverse effects of telomere loss and cooperates with telomere dysfunction to accelerate carcinogenesis. Cell 1999, 97, 527-538. 35.Ely S.A., et al.: Telomerase activity in B-cell non-Hodgkin lymphoma. Cancer 2000, 89, 445-452. 36.MacNamara B., et al.: Telomerase activity in relation to pro- and anti-apoptotic protein expression in high-grade non-Hodgkin´s lymphomas. Haematologica 2001, 86, 386-393. 37.Norrback K.F., et al.: Telomerase activation in normal B lymphocytes and non-Hodgkin´s lymphomas. Blood 1996, 88, 222-9. 38.Norrback K.F.: Telomeres and telomerase in normal and malignant hematopoietic cells. Thesis Umea University 2000. 39.Ohyashiki J.H., et al.: Telomeres and telomerase in hematologic neoplasia. Oncogene 2002, 21, 680-687. 40.Xu D., et al.: Telomerase activity in plasma cell dyscrasias. Br. J. Cancer 2001, 84, 621-625.