© Borgis - Nowa Stomatologia 4/2009, s. 117-121
*Sylwia Chroma, Ewa Iwanicka-Grzegorek
Antybakteryjne peptydy ślinowe – na podstawie piśmiennictwa
Salivary antimicrobial peptide – literature review
Zakład Stomatologii Zachowawczej Instytutu Stomatologii Warszawskiego Uniwersytetu Medycznego
Kierownik Zakładu: prof. dr hab. n. med Elżbieta Jodkowska

Wstęp
Skomplikowana struktura skóry tworzy doskonałą ochronę mechaniczną i biologiczną przed destrukcyjnym działaniem środowiska zewnętrznego, w tym przed mikrobiologiczną inwazją bakteryjną i grzybicami. Nieliczne otwory, w tym jama ustna pozbawione tej naturalnej mechanicznej ochrony wytworzyły bardzo skuteczną barierę biologiczną. Przed niekontrolowaną inwazją bakterii i grzybów jamę ustną zdrowego człowieka chroni nabłonek błony śluzowej, ślina, leukocyty. W ślinie zdrowego człowieka występuje szereg substancji, których zadaniem jest redukcja patogenów są to: immunoglobuliny (IgA, IgG), laktoferryna, histatyny, aglutyniny oraz niedawno opisane peptydy o działaniu przeciwbakteyjnym, w tym, defenzyny alfa i beta, LL-37, katelicyny oraz adrenomedullina. (1, 2)
Endogenne peptydy przeciwbakteryjne
Peptydy o działaniu przeciwbakteryjnym występują w całym organizmie, jednakże szczególnie w miejscach narażonych na infekcję. Peptydy o działaniu antybakteryjnym stanowią nieodłączny element systemu odpornościowego gospodarza. Substancje te produkują komórki ludzkiego nabłonka pokrywającego różne narządy. Peptydy przeciwbakteryjne mogą występować jako stały komponent osłony zdrowego organizmu, ale również infekcja i/lub uszkodzenia tkanek powodują wzrost syntezy i wydzielania różnych peptydów przeciwbakteryjnych. Obecnie zidentyfikowano szeroką gamę endogennych peptydów o działaniu przeciwbakteryjnym. Najlepiej poznanymi i zbadanymi są dwie grupy peptydów nazywanych defenzynami: alfa i beta, które różnią się między sobą specyficznością działania. Stwierdzono, że peptyd z grupy beta-defenzyn (hBD-1) jest stale produkowany w nabłonku zdrowego organizmu, natomiast analogi hBD-2 and -3 są indukowane infekcją. (3, 4, 5, 6)
Beta-defenzyny zostały po raz pierwszy opisane w roku 1995 przez Zasloff. Tworzą one grupę peptydów dodatnio naładowanych, zawierają trzy wewnątrz cząsteczkowe mosty dwusiarkowe, stabilizujące strukturę (ryc. 1).
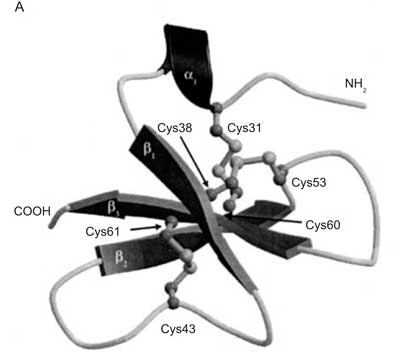
Ryc. 1. Schemat struktury beta defenzyny (hBD-2) [F Bauer, K Schweimer, E Kluver, J-R Conejo-Garcia, W-G Forssmann, P Rosch, K Adermann, H Sticht Structure determination of human and murine_beta-defensins reveals structural conservation in the absence of significant sequence similarity Protein Sci.2001 10: 2470-2479].
Beta-defenzyna (hBD-1) jest białkiem aktywnym wobec bakterii Gram(-). Substancją szczególnie aktywną wobec bakterii Gram(-), Escherichia coli, Pseudomonas aeruginosa, drożdży ( Candida tropicalis, Candida albicans) jest beta-defenzyna hBD-2, która wykazuje również bakteriostatyczny efekt na Staphylococcus aureus.
Alfa-defenzyny są grupą białek nieefektywnych wobec większości bakterii z wyjątkiem Capnocytophaga ochracea i drożdży Candida albicans. W badaniach doświadczalnych obserwowano wzrost alfa defenzyn szczególnie w infekcjach drożdżakowych (7). Wspólnie z białkiem LL-37 działają bakteriobójczo w stosunku do Escherichia coli i Staphylococcus aureus.
Beta-defenzyny hamują replikację wirusa HIV. W badaniach szwedzkich naukowców nad wpływem antybakteryjnych peptydów na aktywność wirusa HIV wykryto, że białka z grupy alfa-defenzyn (hBD-1 wraz z hBD-2) oraz LL-37 wpływają hamująco na rozwój wirusa HIV. Wzrost stężenia peptydów o właściwościach antybakteryjnych jest indukowany w organizmie na skutek nadkażenia bakteryjnego w AIDS. Zwiększone stężenie peptydów przeciwbakteryjnych ma jednocześnie działanie hamujące rozwój wirusa HIV. (8) Inne doświadczenia wykazały, że hBD2 posiadając właściwości redukujące stan zapalny wzmaga odpowiedź gospodarza, polegającą na wzroście aktywności naczyniotwórczej organizmu. (9)
Działanie defenzyn również obserwowano w powikłaniach cukrzycowych. Cukrzyca jest chorobą bardzo często spotykaną w społeczeństwie. Powikłaniem w zaawansowanej cukrzycy jest powstawanie licznych trudno gojących się ran głównie na stopach. Na modelu zwierzęcym zobrazowano wpływ hBD-3 na redukcję bakterii głównie Staphylococcus aureus kolonizujących rany osób chorych na cukrzycę. Wykazano, że hBD3 znacznie redukuje ilość bakterii i wpływa na proces gojenia ran. (10)
Peptydy przeciwbakteryjne w jamie ustnej
W jamie ustnej beta-defenzyny można spotkać w nabłonku pokrywającym język, dziąsła, przewodach wyprowadzających gruczołów ślinowych, błonie śluzowej jamy ustnej.
Beta-defenzyny występujące w jamie ustnej, produkowane są zarówno w zdrowym, jak i zainfekowanym nabłonku, natomiast w nabłonkach pokrywających inne narządy beta-defenzyny produkowane są tylko w odpowiedzi na działanie patogenów. (1, 3, 4, 7, 12) (ryc. 2)
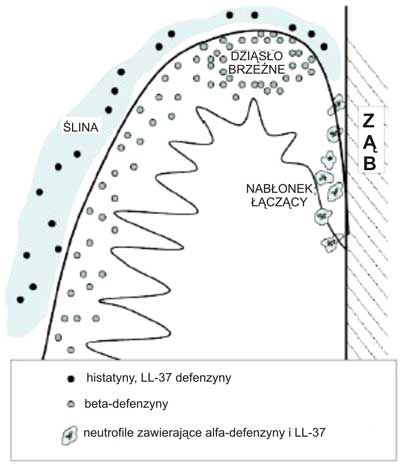
Ryc. 2. Lokalizacja rozmieszczenia antybakteryjnych peptydów w dziąśle. Rycina modyfikowana na podstawie [BA. Dale, LP Fredericks Antimicrobial Peptides in the Oral Environment: Expression and Function in Health and Disease. Curr Issues Mol Biol. 2005; 7: 119-133.]
Bardzo istotną właściwością beta-defenzyny jest brak aktywności bakteriobójczej wobec bakterii komensalnych bytujących w jamie ustnej. Peptyd hBD-1 jest produkowany przez ślinianki, wzrost produkcji tej substancji w okresie infekcji chroni ślinianki przed wsteczną infekcją bakteryjną. (11) Beta defenzynę hBD-1 mogą wydzielać także komórki nabłonka dziąsła, języka, policzka. (12) Ludzka hBD-1 ma silne właściwości przeciwgrzybiczne. Zbadano hipotezę pomiędzy zależnością obecności zmian w genie kodującym hBD-1 a infekcjami oportunistycznymi Candida albicans. Wykazano, że obecność sześciu polimorficznych nukleotydów w genie DEFB1 kodującym hBD-1 ma związek z występowaniem kandydozy. Wykazano związek pomiędzy SNP w piątym nie podlegającym translacji regionie (C-G w pozycji 44) a grzybicą. Osoby, które miały SNP allele 2(G), miały w większości niski poziom Candida w ślinie. Badania wykazują, że genetyczne zmiany w genie DEFB-1 kodującym hBD-1 przyczyniają się do większej skłonności do występowania infekcji Candida w jamie ustnej. (13) Badano też ocenę aktywności antybakteryjnej śliny u osób zdrowych i z kandydozą, stwierdzono, że u osób chorych z uwagi na dużą zawartość peptydów antybakteryjnych ślina ma większą aktywność unieszkodliwiania bakterii, niż ślina osób zdrowych. (14)
Sekrecja beta-defenzyny hBD-2 występuje w zdrowym nabłonku, gdzie jest indukowana przez bakterie komensalne, a także w nabłonku zainfekowanym, w którym za wzrost sekrecji odpowiedzialne są patogeny, zwłaszcza Prophyromonas gingivalis bytujący w jamie ustnej.
Wydzielanie peptydów antybakteryjnych związane jest z działalnością patogenów i produkcją przez nie substancji, wyzwalających obecność TNF alfa, iL1 beta, iL-8, będących czynnikami indukującymi wyzwalanie hBD-2.
Wydzielanie hBD-2 polega na przekazaniu sygnału do komórki nabłonka przez receptory TLR. Pochodzące z rozpadu bakterii Gram(-) kwasy lipolityczne i peptydoglikany sygnalizują przez TLR 2 i TLR 4 aktywację kaskady NFkappaB i ścieżek JNK. Jest to przykład aktywacji przez czynniki patogenne. W inny sposób aktywują hBD-2 bakterie komensale bytujące w jamie ustnej, wykorzystując JNK i p 38 kinazę proteinową aktywowaną miogenem (MAPK) – czyli ścieżki skojarzone z odpowiedzią na cytokiny i na stres. Natomiast w nabłonkach pokrywających inne narządy aktywacja hBD-2 odbywa się tylko poprzez kaskadę NF kappaB (1, 15). Widoczne dwa różne sposoby aktywacji hBD-2, związane są ze specyfiką środowiska jamy ustnej i działaniem ochronnym na bakterie komensalne, są pierwszą barierą chroniącą ten obszar przewodu pokarmowego człowieka przed patogenami.
Receptory TLR 2 i 4 regulowane są przez IF gamma, ale bakteryjne LPS i CD14 są kofaktorem w połączeniu bakteryjnych LPS do TLR 4.
Drugą grupę receptorów stanowią receptory PAR będące szczególnie wrażliwe na działanie proteaz, występujących u bakterii odpowiedzialnych za powstawanie chorób przyzębia, takich jak: Prophyromonas gingivalis, Bacteroides forsytus i Treponema denticola. Proteazy są częścią mechanizmu wirulencyjnego tych trzech szczególnie patogennych bakterii. Reakcja proteaz z receptorem PAR pozwala prawidłowo odpowiedzieć na bytujące w czasie infekcji patogeny, natomiast komórka nie odpowiada na sygnały pochodzące od bakterii komensalnych, które nie posiadają proteaz.
Szerokie spektrum działania wobec bakterii Gram (-) i Gram(+) oraz grzybów jest cechą charakterystyczną kolejnej beta-defenzyny hBD-3. Wydzielanie hBD-3 stymulowane jest przez IF gamma oraz Actinobacillus actinomycetemcommitans, który jest bakterią odpowiedzialną za progresję chorób przyzębia. Obie beta-defenzyny (hBD-2 i hBD-3) wydzielane są przez warstwę kolczystą nabłonka a największa ekspresja tych substancji występuje na brzegu dziąsłowym, który jest miejscem kontaktu dziąsła z płytką bakteryjną. Nie obserwuje się podklas tych beta-defenzyn w nabłonku łączącym kieszonki dziąsłowej.
Kolejna, niedawno opisana, beta-defenzyna hBD-4, jest odpowiedzialna za hamowanie wzrostu bakterii Gram(-) i Gram(+), Pseudomonas aeruginosa oraz grzybów.
Innymi peptydami antybakteryjnym występującymi w ślinie są kalprotektyny. Ich heterodimeryczna cząsteczka zawiera wapń i cynk.
Powyżej zamieściliśmy fragment artykułu, do którego możesz uzyskać pełny dostęp.
Mam kod dostępu
- Aby uzyskać płatny dostęp do pełnej treści powyższego artykułu albo wszystkich artykułów (w zależności od wybranej opcji), należy wprowadzić kod.
- Wprowadzając kod, akceptują Państwo treść Regulaminu oraz potwierdzają zapoznanie się z nim.
- Aby kupić kod proszę skorzystać z jednej z poniższych opcji.
Opcja #1
29 zł
Wybieram
- dostęp do tego artykułu
- dostęp na 7 dni
uzyskany kod musi być wprowadzony na stronie artykułu, do którego został wykupiony
Opcja #2
69 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 30 dni
- najpopularniejsza opcja
Opcja #3
129 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 90 dni
- oszczędzasz 78 zł
Piśmiennictwo
1. M Mathews et al.: Production of Beta –Defensin Antimicrobial Peptides by the Oral Mucosa and Salivary Glands. Infection and Immunity 1999; 2740-2745. 2. C Shelburne et al.: Induction of Beta – Defensin Resistance in the Oral Anaerobe Prophyromonas gingivalis. Antimicrobial Agents and Chemiotherapy 2005; 183-187. 3. Harder J et al.: Isolation and characterization of human beta -defensin-3, a novel human inducible peptide antibiotic. J Biol Chem 2001; 276: 5707-13. 4. Bowdish DM, Davidson DJ, Hancock RE: Immunomodulatory properties of defensins and cathelicidins. Curr Top Microbiol Immunol 2006; 306: 27-66. 4. 5. Krisanaprakornkit S et al.: Expression of the peptide antibiotic human beta-defensin 1 in cultured gingival epithelial cells and gingival tissue. Infect Immun 1998; 66: 4222-8. 6. Garcia JR et al.: Identification of a novel, multifunctional beta-defensin (human beta-defensin 3) with specific antimicrobial activity: its interaction with plasma membranes of Xenopus oocytes and the induction of macrophage chemoattraction. Cell Tissue Res 2001; 306: 257-64. 7. Sawaki K et al.: Immunohistochemical study on expression of alpha defensin and beta-defensin -2 in human buccal epithelia with candidiasis. Oral Dis 2002; 8(1): 37-41. 8. Bose SU et al.: Donald CDPax 2 oncogene negatively regulates the expression of the host defense peptide human beta-defensin -1 in prostate cancer. Mol Immunol 2009; 46 (6): 1140-8. 9. Hirsch T et al.: Human beta-defensin -3 promotes wound healing in infecteddiabetic wounds. J Gene Med 2009; 11(3): 220-8. 10. Liu HW, He GX, Sun H: Expression and significance of human beta-defensin in recurrent nasal polyps. 11. Sahasrabudhe KS et al.: Expression of the antimicrobial peptide, human beta-defensin 1, in duct cells of minor salivary glands and detection in saliva. Dent Res 2000; 79(9): 1669-74. 12. Mathews M et al.: Production of beta-defensin antimicrobial peptides by the oral mucosa and salivary glands. Infect Immun1999; 67(6): 2740-5. 13. Jurevic RJ et al.: Single-nucleotide polymorphisms (SNPs) in human beta-defensin 1: high-throughput SNP assays and association with Candida carriage in type I diabetics and nondiabetic controls. J Clin Microbiol 2003; 41(1): 90- 6. 14. Tanida T et al.: Decreased excretion of antimicrobial proteins and peptides In saliva of patients with oral candidiasis. J Oral Pathol Med. 2003; 32(10): 586-94. 15. Diamond G, Beckloff N, Ryan LK: Host defense peptides in the ora cavity and the lung: similarities and differences. J Dent Res 2008; 87(10): 915-27. 16. De Smet K, Contreras R: Human antimicrobial peptides: defensins, cathelicidins and histatins” Biotechnol Lett.2005; 27(18): 1337-47. 17. Dale Ba et al.: Oral antimicrobial peptides and biological control of caries” BMC Oral Health 2006; 15(6): 13. 18. Abiko Y, Nishimura M, Kaku T: Defensins in saliva and the salivary glands. Med Electron Microsc 2003; 36 (4): 247-52. 19. Yin C et al.: Mouse salivary glands and human beta-defensin-2 as a study model for antimicrobial gene therapy: technical considerations. Int J Antimicrob Agents 2006; 28(4): 352-60 20. Ouhara K et al.: Susceptibilities of periodontopathogenic and cariogenic bacteria to antibacterial peptides {beta}- defensins and LL37, produced by human epithelial cells. J Antimicrob Chemother 2005; 55(6): 888-96. 21. Ji S et al.: Toll-like receptor 2 and NALP 2 mediate induction of human beta-defensins by fusobacterium nucleatum in gingival epithelial cells” Infect Immun.2009; 77(3): 1044-52. 22. Tao R et al.: Salivary antimicrobial peptide expression and dental caries experience in children. Antimicrob Agents Chemother 2005; 49(9): 3883-8. 23. Gosh SK et al.: Quantification of human beta-defensin -2 and -3 in body fluids:application for studies of innate immunity. Clin Chem. 2007; 53(4): 757-65. 24. Maisetta G et al.: Susceptibility of Streptococcus mutans and Actinobacillus actinomycetemcomitans to bacterial activity of human beta-defensin 3 in biological fluids. Antimicrob Agents Chemother 2005; 49(3): 1245-8. 25. Shi Y et al.: Disinfection of maxillofacial silicone elastomer using a novel antimicrobial agent: recombinant human beta-defensin -3. Eur J Clin Microbiol Infect Dis 2008; 8. 26. Juusola J, Ballantyne J: Multiplex m RNA profiling for the identification of body fluids” Forensic Sci Int 2005; 152(1): 1-12. 27. Varoga D et al.: The role of human beta-defensin -2 in bone. J Anat 2008; 213(6): 749-57.
