© Borgis - Medycyna Rodzinna 2/2008, s. 34-47
Justyna Kosińska1, Katarzyna Billing-Marczak1, *Marcin Krotkiewski2
Nowe nieznane funkcje witaminy D
NEW UNKNOWN FUNCTIONS OF VITAMIN D
1 Dział Badań i Rozwoju Nowych Produktów, Biotech-Varsovia Sp. z o.o. – Sp. K. w Warszawie Dyrektor Działu: dr n. biol. Katarzyna Billing-Marczak 2 The Sahlgrenska Academy at Göteborg University, Institute of Clinical Neuroscience, Göteborg, Sweden
Summary
Vitamin D together with vitamin K contribute to the absorption of calcium and mineralization of bones. Recent epidemiological studies are indicating several nonskeletal effects of vitamin D. Negative correlations with vitamin D serum levels were found for hypertension, diabetes type 1 and 2, obesity, metabolic syndrome, chronic inflammatory conditions and different forms of cancers. In particular incidence of colon and anal cancer, cancer of prostate, ovarian and uterus cancer as well as breast cancer and Hodgkin lymphoma is negatively correlated with serum concentration of vitamin D. Multiorgans effects of vitamin D depend on the presence in different tissues of specific receptors (VDR). Vitamin D3 combines with VDR and subsequently with RXR (Retinoid X Receptor). The complex vitamin D – VDR – RXR is later functioning as typical transcription factor activating specific sequences in promoters of several genes – VDRE (Vitamin D Response Elements) – expressing different proteins. This mechanism responds for the increased synthesis of and sensitivity to insulin. Vitamin D is inhibiting the activity of transcription factor NFkB and decreasing the production of proinflammatory cytokines. The immunomodulating effect of vitamin D is also expressed as lower incidence of virus and bacterial infections. Vitamin D is slowing down the shortening of telomeres indicating possible influence on aging and longevity. Supplementation with vitamin D is highly recommended particularly for older people. Recommended daily doses should be strongly increased.

Źródła witaminy D
Witamina D występuje w kilku formach. Wyróżnić możemy witaminę D1 (kalcyferol), witaminę D2 (ergokalcyferol) oraz witaminę D3 (cholekalcyferol). Witamina D1 występuje głównie w tranie, witamina D2 – w roślinach, zaś witamina D3 powstaje w tkance skórnej ludzi i zwierząt.
Efekty metaboliczne witaminy D2 i witaminy D3 są podobne, jednak wiążą się one z różnymi typami białek osocza, w efekcie czego witamina D3 działa dłużej i jest 2-10 razy bardziej efektywna niż witamina D2 (1, 2).
Witamina D3 jest jedną z niewielu witamin, którą organizm może wyprodukować endogennie. Potrzebne do tego procesu promieniowanie ultrafioletowe (UVB – frakcja B tego promieniowania) umożliwia przemianę zawartego w skórze 7-dehydrocholesterolu (tzw. prowitamina D3) w cholekalcyferol (witamina D3) (3). Jednak synteza wystarczającej ilości witaminy D drogą fotolizy pochodnej cholesterolu jest ograniczona przez działanie takich czynników jak czynniki geograficzne i środowiskowe (szerokość geograficzna, pora roku, warunki atmosferyczne) oraz inne czynniki (np. ubranie), które ograniczają dopływ promieni słonecznych do powierzchni skóry (4). Jakiekolwiek zmiany w ilości docierających do nas fotonów UVB, mogą mieć z kolei dramatyczny wpływ na skórną produkcję witaminy D. Badania Webb´a i innych współautorów wykazały, że intensywność syntezy witaminy D w skórze w okresie zimy, jak również powyżej i poniżej 35°N i 35°S szerokości geograficznej, jest prawie niewykrywalna (5, 6, 7). W Polsce synteza witaminy D pod wpływem działania promieni UVB zachodzi praktycznie jedynie przez parę godzin w ciągu dnia w okresie letnim.
Do czynników biologicznych wpływających na szybkość syntezy witaminy D zalicza się wiek i zawartość melaniny w skórze (4). Według Clemensa i wsp. zwiększona pigmentacja skóry może nawet od 5 do 10 razy zmniejszać skórną syntezę witaminy D (6, 8, 9). Osoby o jasnej karnacji szybciej przekształcają 7-dehydrocholesterol niż osoby o karnacji ciemnej. Badania wykazały, że przy jasnej karnacji wystarczy 20-minutowa ekspozycja na promieniowanie ultrafioletowe (aż do lekkiego zaczerwienienia) w celu wytworzenia około 250 μg witaminy D (około 10 000 JE, gdyż 10 μg = 400 JE), podczas gdy zalecana dzienna dawka wynosi około 75 ?g. Osoby o ciemniejszej karnacji potrzebują około 5-10 razy więcej czasu, żeby doszło do syntezy podobnej ilości witaminy, jak u osób o karnacji jasnej (6, 8, 9). Ponadto, Matsuoka i wsp. donoszą, że stosowanie kremów z filtrem może również redukować penetrację UVB do naskórka nawet o 97,5% wpływając niekorzystnie na proces syntezy witaminy D (6).
Z innych badań wynika, że intensywność skórnej syntezy witaminy D ulega stopniowej redukcji wraz z wiekiem. Z badań porównawczych wynika, że po ekspozycji na promienie UVB, ilość powstałej w skórze witaminy D u osób młodszych była co najmniej dwa razy większa niż u osób starszych. Potwierdzają to obserwacje, według których ilość obecnego w skórze 7-dehydrocholesterolu jest negatywnie skorelowana z wiekiem (10).
Ponadto, ilość powstającej witaminy D zależy od powierzchni eksponowanej skóry. Im większa powierzchnia skóry wystawiona na działanie promieni UVB, tym więcej witaminy D powstaje w jednostce czasu. Proces ten ulega jednak szybkiemu wysyceniu i po około 30 minutach ulega zahamowaniu (7, 11). Wynika to z faktu, że tylko około 15% obecnego w skórze 7-dehydrocholesterolu ulega przekształceniu w witaminę D3 podczas początkowych 30 minut ekspozycji na światło słoneczne (12).
Mimo że promieniowanie ultrafioletowe jest głównym źródłem witaminy D, istotne znaczenie ma również żywność. Istnieje tylko kilka naturalnych źródeł witaminy D (3), do których zaliczamy m.in. mleko, mleko sojowe, masło (13), ryby i niektóre grzyby (np. kurki) (14). Natomiast większość produktów to żywność wzbogacana w ten składnik. W Szwecji, wiele artykułów spożywczych i suplementów diety zawiera w swoim składzie dodatek witaminy D (3). Średnia ilość witaminy D dostarczanej w diecie w krajach europejskich waha się w granicach 2,5-4,0 μg/dzień. W wielu krajach rekomendowane jest dostarczenie do 10 μg/dzień do 2. roku życia, zaś dla kobiet w ciąży i dla osób starszych – 7,5 μg/dzień (15, 16, 17, 18).
Mimo, iż wiele pokarmów jest wzbogaconych w witaminę D, jej niedobór staje się coraz bardziej powszechny. Unikanie promieni słonecznych, stosowanie filtrów ochronnych, szerokość geograficzna, ubrania i zwyczaje kulturowe dodatkowo pogłębiają problem związany z jej deficytem. Wśród Innuitów na Grenlandii, którzy przeszli na dietę europejską, jak również wśród emigrantów z Afryki, którzy po przybyciu do Europy kontynuują dietę ubogą w ten składnik oraz ubierają się zasłaniając twarz (w przypadku kobiet), niedobór witaminy D staje się coraz bardziej zauważalny (3).
Metabolizm witaminy D
Proces aktywacji witaminy D przebiega wieloetapowo. Witamina D magazynowana jest w tkance tłuszczowej oraz w wątrobie. W tej ostatniej ulega przekształceniu do 25-hydroksywitaminy D (kalcidiol, kalcifediol, 25OHD) w wyniku hydroksylacji atomu węgla C25. Badania wykazały, że proces ten zachodzi również w komórkach nabłonka – keratynocytach (3). W kolejnym etapie metabolicznej przemiany, kalcidiol ulega hydroksylacji pod wpływem 1-α-hydroksylazy do 1,25-dwuhydroksywitaminy D (kalcitriol, 1,25(OH)2D) – najbardziej aktywnej formy witaminy D (formy hormonalnie aktywnej). Pierwotnie sądzono, że proces ten zlokalizowany jest tylko w nerkach. Obecnie wiadomo, że przejście 25OHD do 1,25(OH)2D ma miejsce także w przewodzie pokarmowym, układzie kostnym, mięśniach, neuronach, trzustce, gruczole krokowym i limfocytach (19). W przypadku zaburzeń w funkcjonowaniu nerek poziom kalcitriolu w układzie krążenia spada jednak do wartości niewykrywalnych. Wynika z tego, że jeśli inne tkanki i organy produkują hormonalnie aktywną formę witaminy D, to najwidoczniej nie jest ona uwalniania do krwioobiegu, lecz działa lokalnie, w obrębie komórki, w której powstała lub w komórkach sąsiadujących. Może to świadczyć o parakrynnej i autokrynnej regulacji systemu wydzielania witaminy D.
Kalcitriol jest najbardziej aktywnym hormonem sterydowym, działającym już w stężeniach pikogramowych. Jego stężenie w surowicy krwi podlega regulacji za pośrednictwem nerek oraz przytarczyc i jest zależne od poziomu wapnia w osoczu (3).
Cytochromy z rodziny P 450 (CYP-24 i CYP-3A4) oraz niektóre leki (fenobarbital, fenytoina, rifampicyna) odpowiadają za inaktywację hormonalnej formy witaminy D. Konsekwencją ich działania jest deficyt kalcitriolu i wzrost ryzyka rozwoju chorób, takich jak osteomalacja (20).
Aktywność witaminy D – receptory
Witamina D działa za pośrednictwem specyficznych receptorów o charakterze typowych czynników transkrypcyjnych (3). Spośród wszystkich postaci witaminy D, tylko kalcitriol jako forma metabolicznie aktywna funkcjonuje poprzez aktywację receptora VDR (ang. Vitamin D Receptor) (21). Receptor ten należy do rodziny jądrowych receptorów specyficznych dla hormonów steroidowych, do której zalicza się również receptory hormonów tarczycy, receptor retinowy RXR (ang. Retinoic X Receptor) oraz receptor aktywowany przez czynniki proliferacji peroksysomów PPAR (ang. Peroxisome Proliferator – Activated Receptor). Znajdujący się na ludzkim chromosomie 12cen–q12 gen receptora VDR, jest miejscem szeregu polimorfizmów (najczęściej występującym polimorfizmem jest (A)n – Repeated Single Nucleotide Polymorphism) (21), powiązanych z pierwotną nadczynnością przytarczyc, gęstością układu kostnego i wysokością ciała (22). Ponadto, badania wykazały, że gen kodujący VDR występuje z reguły w pobliżu genu decydującego o ekspresji 1-α-hydroksylazy (21).
Związanie się 1,25(OH)2D do VDR prowadzi do transkrypcji genów podlegających regulacji przez kalcitriol (ryc. 1). Jest to proces wieloetapowy i rozpoczyna się od utworzenia kompleksu 1,25(OH)2D z VDR, który wiąże się następnie z receptorem retinowym RXR. Heterodimer VDR–RXR oddziałuje za pośrednictwem 1,25(OH)2D ze specyficznymi sekwencjami DNA w promotorach genów regulatorowych, czyli z tzw. VDRE (ang. Vitamin D Response Elements). Do powstałego kompleksu przyłączają się czynniki transkrypcyjne powodując zmianę szybkości transkrypcji genów. Regulacja za pośrednictwem kalcitriolu dotyczy szczególnie genów kodujących cytokiny, genów odpowiedzialnych za produkcję białek niezbędnych do prawidłowego funkcjonowania komórek β trzustki (19) oraz genów kodujących: białka wiążące wapń (CaBP, ang. Calcium-Binding Protein), 24-hydroksylazę, antygen specyficzny dla gruczołu krokowego (PSA, ang. Prostate-Specific Antigen), parathormon (PTH), osteokalcynę, osteopontynę oraz kolagen (23).
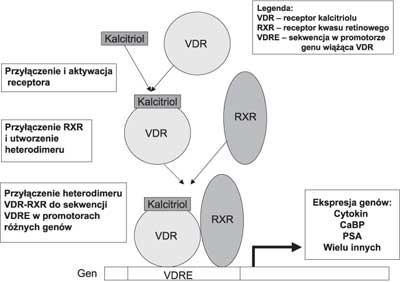
Ryc. 1. Regulacja ekspresji genów za pośrednictwem kalcitriolu (opis w tekście).
Obecność receptorów VDR stwierdzono nie tylko w organach zaangażowanych w utrzymanie homeostazy mineralnej, jak jelito, kość, nerka, przytarczyce, lecz również w tkankach i organach innego rodzaju (24, 25) (ryc. 2). Jednak efekty kalcitriolu w stosunku do innych tkanek niż tkanka kostna wymagają stężeń rzędu 10-10 mol/l, które to znacznie przewyższają (100-1000 razy) poziom 1,25(OH)2D potrzebny do utrzymania homeostazy wapnia. Biorąc pod uwagę fakt, iż niektóre komórki układu immunologicznego, takie jak makrofagi, wydzielają 1,25(OH)2D wiemy, że wysokie stężenia kalcitriolu mogą być osiągane lokalnie, np. w obszarach objętych stanem zapalnym. Jednak podawanie większych dawek 1,25(OH)2D w celach innych niż wpływ na tkankę kostną (np. w celach immunomodulacyjnych, przeciwzapalnych, antyproliferacyjnych) mogłoby doprowadzić do niebezpiecznej hiperkalcemii. Prowadzone aktualnie badania nad analogami witaminy D mogą rozwiązać ten problem. Związki te bowiem zostały tak zmodyfikowane, że ich wpływ na homeostazę wapniową jest znacznie mniejszy niż kalcitriolu. Dzięki temu, można je będzie podawać w większych dawkach bez ryzyka wystąpienia hiperkalcemii (19).

Ryc. 2. Wpływ witaminy D na organizm człowieka. Syntetyzowana w skórze witamina D trafia do krwioobiegu, skąd przenoszona jest do różnych tkanek i organów. Obecność receptorów VDR w organach zaangażowanych w utrzymanie homeostazy mineralnej (jelito, kość, nerka, przytarczyce), jak również w tkankach i organach innego rodzaju (komórki β trzustki, komórki układu immunologicznego) świadczy o szerokim spektrum działania kalcitriolu (opis w tekście).
Wpływ witaminy D na układ kostny
Witamina D odgrywa istotną rolę w procesie wchłaniania wapnia w jelicie cienkim (ryc. 2). Jej niedobór prowadzi do krzywicy, osteomalacji (odpowiednik krzywicy u dorosłych) oraz osteoporozy. O funkcji witaminy D w procesie mineralizacji kości może np. świadczyć fakt, iż stan tkanki kostnej ulega pogorszeniu w okresie zima/wiosna, kiedy synteza witaminy D pod wpływem działania promieni słonecznych jest znacznie ograniczona.
Poziom 25OHD w surowicy krwi jest pozytywnie skorelowany ze stopniem mineralizacji kości, zaś negatywnie z koncentracją parathormonu (PTH) (ryc. 2). Przewlekły niedobór witaminy D prowadzi do zaburzenia procesu wchłaniania wapnia w jelicie cienkim w rezultacie czego, stężenie wapnia we krwi spada. W odpowiedzi na nieprawidłowy poziom wapnia we krwi, przytarczyce zwiększają produkcję PTH, który kosztem wapnia zgromadzonego w kościach, uzupełnia jego niedobory w surowicy krwi. Proces ten przebiega poprzez aktywację osteoklastów, co skutkuje destrukcją tkanki kostnej (26).
Badania naukowe dowodzą, że suplementacja kalcidiolem prowadząca do wzrostu surowiczego poziomu witaminy D do około 71-99 nmol/l, znacznie zmniejszyła ryzyko złamań kości u szczurów. Z kolei podaż witaminy D w ilości powodującej wzrost jej stężenia w surowicy krwi do wartości około 54-62 nmol/l, nie przyniosła podobnych rezultatów. Zdania co do optymalnego stężenia witaminy D w surowicy krwi są podzielone. Dawson-Hughes i wsp. postulują, że średni poziom witaminy D w surowicy krwi, zmniejszający ryzyko złamań kości, powinien wynosić około 75 nmol/l. Inni badacze zakładają, że właściwymi wartościami prewencyjnymi są 50-80 nmol/l lub 70-80 nmol/l. Ponieważ pozytywny efekt wywołała suplementacja witaminą D w ilości 17,5-20,5 ?g na dobę, przypuszcza się, że jej średnia optymalna dobowa dawka to około 20 ?g na dzień. Wiele z tych badań opartych zostało jednak na leczeniu osteoporozy kombinacją witaminy D w połączeniu z wapniem, a nie samą witaminą D (26). Wpływ większych dawek witaminy D nie został dostatecznie przebadany, jednakże kontrolowane studium kliniczne u starszych kobiet wykazało, że podawanie witaminy D2 w ilości 54 ?g na dzień, znamiennie zwiększyło masę części rdzeniowej kości śródręcza (26).
Istotny wpływ witaminy D na stopień mineralizacji kośćca potwierdzają również badania, według których samo leczenie osteoporozy przy użyciu bisfosfonianów nie zawsze jest wystarczające do uzyskania optymalnych efektów terapeutycznych. Heckman i wsp. wykazali, że podawanie witaminy D pacjentom, którzy nie reagowali na terapię bisfosfonianami, zaowocowało wzrostem gęstości masy kostnej o 1,45% w przypadku kości lędźwiowej i o 1,15% w przypadku szyjki udowej (27). Być może działanie bisfosfonianów uzależnione jest od odpowiedniego poziomu witaminy D w ustroju. Także przewlekłe degeneracyjne zapalenie stawów (artroza) ma statystycznie lepsze rokowania przy wyższej suplementacji witaminą D (powyżej 75 nmol/l w surowicy krwi) (28).
Wpływ witaminy D na układ mięśniowy
Szereg doniesień naukowych podaje związek między niedoborem witaminy D a schorzeniami tkanki mięśniowej, takimi jak miopatia. Zauważono, że deficyt witaminy D sprzyja zmniejszeniu ilości białych włókien mięśniowych (typ II), co jest typowym objawem procesów starzenia. Na poziomie molekularno-biologicznym witamina D wpływa na transport komórkowy jonów wapnia oraz syntezę niezbędnych dla prawidłowego funkcjonowania mięśni białek, takich jak aktyna i troponina. Dowiedziono, że prawidłowe funkcjonowanie mięśni jest pozytywnie skorelowane z poziomem 25OHD, zaś negatywnie z PTH. Nie wykazano natomiast żadnego wpływu 1,25(OH)2D na siłę mięśni. Suplementacja witaminą D pozwala odwrócić atrofię mięśniową. Badania wykazały, że pierwsze rezultaty widoczne były już po 3 miesiącach kuracji witaminą D, jednakże najlepsze efekty obserwowano po 6-12 miesiącach leczenia (29).
Naukowcy zwrócili również uwagę na obecność polimorfizmów w obrębie genu kodującego receptor VDR jako potencjalnej przyczyny zaburzeń w prawidłowym funkcjonowaniu mięśni. Do polimorfizmów tych zaliczamy FokI oparty na tranzycji T–>C w obrębie egzonu 2 genu kodującego VDR [30] oraz BsmI będący fragmentem restrykcyjnym na końcu 3´ genu kodującego receptor VDR (31).
Hopkinson i wsp. zbadali wpływ różnych genotypów receptora VDR na siłę mięśnia czworogłowego uda u pacjentów cierpiących na przewlekłą obturacyjną chorobę płuc (ang. COPD; Chronic Obstructive Pulmonary Disease) biorąc pod uwagę fakt, iż zaburzenia w funkcjonowaniu i morfologii wspomnianego mięśnia należą do typowych powikłań COPD. Naukowcy zaobserwowali, że chorzy charakteryzowali się znacznie słabszą siłą mięśni szkieletowych w porównaniu z grupą kontrolną. Jednocześnie zauważyli, że homozygoty pod względem allelu C (tranzycja T–>C) wykazywały mniejszą siłę mięśnia czworogłowego uda niż osoby z genotypami CT lub TT. Zależność ta dotyczyła zarówno pacjentów, jak i osób zdrowych. W przypadku polimorfizmu BsmI zmiany w sile mięśnia czworogłowego zaobserwowano jedynie w grupie z COPD. Homozygoty pod względem allelu b charakteryzowała większa siła mięśni w porównaniu z homozygotami B oraz heterozygotami. Nie odnotowano natomiast wpływu polimorfizmu BsmI na zdolność skurczową mięśni w grupie kontrolnej (32). Co ciekawe, Geusens i wsp. donoszą o większej sile mięśnia czworogłowego u starszych kobiet z allelem b (31) podczas, gdy inny zespół naukowców zaobserwował, że kobiety młodsze z allelem b charakteryzowały się mniejszą siłą ścięgna podkolanowego. Nie zauważono u nich natomiast wpływu allelu b na zdolność skurczową mięśnia czworogłowego uda (33). Z powyższego wynika, że polimorfizmy w obrębie genu kodującego receptor VDR mogą wywierać wpływ na prawidłowe funkcjonowanie mięśni, a efekty ich działania zależą od rodzaju mięśnia oraz wieku.
Na uwagę zasługuje również fakt, iż związanie się witaminy D z jej receptorem prowadzi do proliferacji oraz różnicowania komórek w stronę dojrzałych włókien mięśniowych (34), co zostało potwierdzone badaniami na myszach pozbawionych genu kodującego VDR [35]. Naukowcy donoszą również, że ekspresja genu kodującego VDR maleje z wiekiem, co może tłumaczyć mniejszą siłę mięśni u osób starszych (36).
Z powyższych doniesień wynika, że niedobór witaminy D, różne genotypy receptora VDR, jak i spadek ekspresji kodującego go genu mogą być przyczyną zaburzeń w prawidłowym funkcjonowaniu mięśni oraz tłumaczyć postępujący wraz z wiekiem zanik siły mięśni.
Wpływ witaminy D na ryzyko wystąpienia chorób nowotworowych
Szereg badań epidemiologicznych wykazało, że występowanie najczęstszych form raka, takich jak: rak piersi, rak jelita grubego, rak odbytnicy, rak macicy, rak jajników, rak gruczołu krokowego i białaczka limfatyczna, wykazuje negatywną korelację z poziomem witaminy D w surowicy krwi. Zauważono, że witamina D działa antykancerogennie i znacznie zmniejsza częstość występowania chorób nowotworowych u osób, u których jej poziom jest nie mniejszy niż 30 ng/ml surowicy krwi.
Głównym źródłem witaminy D jest promieniowanie słoneczne. Zaobserwowano, że częstość zachorowań na raka jest większa na obszarach o mniejszym nasłonecznieniu, a mniejsza na terenach, na których światło słoneczne jest dostępne w większych ilościach i przez dłuższy okres czasu. Odwrotną zależność między ekspozycją na promienie UVB a ryzykiem wystąpienia którejś z odmian raka potwierdzają badania epidemiologiczne przeprowadzone w USA, których celem było zbadanie korelacji między śmiertelnością spowodowaną chorobami nowotworowymi a szerokością geograficzną.
Co ciekawe, ryzyko wystąpienia raka jest również zdeterminowane cechami rasowymi. Jak się okazało, ludność rasy czarnej charakteryzuje co najmniej o połowę mniejszy poziom kalcidiolu niż rasę białą. Jednocześnie częstotliwość występowania różnych odmian raka jest u nich większa niż u ludności rasy białej (37). Dyskutuje się, czy różnica w powszechności występowania chorób nowotworowych wśród ludzi różnych ras, może wynikać z różnic szybkości syntezy witaminy D pod wpływem działania promieni UVB, która u osób z ciemniejszą karnacją jest znacznie wolniejsza, niż u osób o karnacji jasnej.
Antykancerogenne działanie witaminy D stwierdzono wobec wielu rodzajów raka. W Szwecji, Smedby i wsp. wykazali, że ekspozycja na promienie UVB zmniejsza ryzyko wystąpienia chłoniaków nieziarniczych (ang. Non Hodgkin Lymphoma; NHL). Osoby, które częściej korzystają z kąpieli słonecznych, rzadziej chorują na ten rodzaj nowotworu niż osoby unikające promieniowania ultrafioletowego (38).
Badania nad grupą pacjentów cierpiących na raka prostaty wykazały, że stężenie we krwi antygenu gruczołu krokowego (ang. Prostate Specific Antygen; PSA), markera przerostu prostaty i raka prostaty, znacznie zmalało po 6-15 miesiącach suplementacji kalcitriolem, w porównaniu z pacjentami otrzymującymi placebo. Przypuszcza się, że witamina D uwrażliwia komórki nowotworowe na cytotoksyczne działanie cytostatyków. Dzięki temu, leczenie raka prostaty tradycyjnymi metodami w połączeniu z suplementacją kalcitriolem, przynosi znacznie lepsze efekty niż poddawanie pacjenta samej chemioterapii (39).
Podobne rezultaty uzyskano w badaniach pilotowych na grupie pacjentów z glejakiem lub astrocytoma. Zaobserwowano, że u trzech na jedenastu badanych, dodatek witaminy D do standardowej chemioterapii, zaowocował znaczną poprawą ich stanu zdrowia (40, 41, 42).
Obecnie wiadomo, że poza tkankami i narządami zaangażowanymi w utrzymanie homeostazy mineralnej, receptory witaminy D występują również w komórkach różnych nowotworów. Obecność receptorów VDR w komórkach czerniaka, nowotworów piersi, jelita grubego i wielu innych odmian raka wskazuje, że funkcje witaminy D nie są ograniczone jedynie do tkanki kostnej, lecz swoim działaniem obejmują również komórki zmienione nowotworowo. Rola witaminy D w profilaktyce i walce z chorobami nowotworowymi wynika z jej działania proróżnicującego i antyproliferacyjnego (3). Witamina D pełni także rolę czynnika hamującego angiogenezę (43) oraz stymulującego apoptozę komórek rakowych (44).
Receptory VDR zlokalizowano m.in. w komórkach nabłonka gruczołu krokowego i okrężnicy. Badania wykazały, że u osób z właściwym poziomem witaminy D w surowicy krwi, ryzyko zachorowania na nowotwory okrężnicy i gruczołu krokowego jest znacznie mniejsze, niż u osób z jej niedoborem. Należy jednak wziąć pod uwagę, że nie tylko poziom samej witaminy D w surowicy krwi ma wpływ na podatność na choroby nowotworowe. Znaczenie ma również polimorfizm obu kopii genu kodującego VDR, który prowadzi do powstania odmiennych genotypów. Od tych ostatnich zależy podatność na różne odmiany raka (37).
Szereg innych badań potwierdziło, że zmniejszony poziom witaminy D zwiększa ryzyko zachorowania na choroby nowotworowe. Wydaje się zatem, że kampania medialna przestrzegająca przed opalaniem ze względu na ryzyko powstania raka skóry, wymaga pewnego zrównoważenia i ostrożności.
Wpływ witaminy D na ryzyko wystąpienia zespołu metabolicznego
W ostatnich latach niedobór witaminy D powiązano z ryzykiem wystąpienia zespołu metabolicznego, skupiającego takie zaburzenia w normalnym funkcjonowaniu organizmu, jak: insulinooporność, nietolerancja glukozy, dyslipidemia, nadciśnienie oraz otyłość typu wisceralnego (jabłkowa) (45), często prowadzące do rozwoju cukrzycy typu 2 oraz zawału mięśnia sercowego (46). Liczne badania wykazały, że ilość spożywanych produktów nabiałowych jest negatywnie skorelowana z masą ciała, nadciśnieniem, homeostazą glukozy i cukrzycą typu 2 (47). Ponieważ witamina D i wapń są głównymi komponentami tego rodzaju żywności, uznano za słuszne stwierdzenie, że zarówno jeden jak i drugi składnik, mogą mieć istotny wpływ na rozwój zespołu metabolicznego.
Zależność między podażą wapnia a ryzykiem wystąpienia otyłości
Davies i wsp. obliczyli, że różnice w podaży wapnia odpowiadają za 3% zmianę wagi ciała (48). Liczne badania naukowe potwierdzają fakt, że dieta bogata w wapń sprzyja zmniejszeniu masy ciała. Mechanizm opiera się prawdopodobnie na wpływie wewnątrzkomórkowych jonów wapnia na adipocyty, komórki tłuszczowe syntetyzujące i magazynujące tłuszcze proste. Przy niskiej podaży wapnia wzrastają poziomy PTH i kalcitriolu, które stymulują uwalnianie jonów Ca2+ w ludzkich adipocytach, aktywując je do wzrostu lipogenezy i inhibicji lipolizy. Proces ten sprzyja wzrostowi masy ciała. Z kolei, wzrost podaży produktów nabiałowych prowadzi do efektu odwrotnego. Obniżone zostają stężenia kalcitriolu i PTH a ilość jonów Ca2+ w adipocytach pozostaje na niskim poziomie, sprzyjając stymulacji lipolizy i inhibicji syntezy oraz magazynowania tłuszczów (49). Można by sądzić, że wapń wpływa na regulację apetytu oraz wydatkowanie energii (50). Jednakże Jacobsen i wsp. doszli do innego wniosku. Według nich dieta bogata w wapń nie wpływa na wydatkowanie energii, czy też procesy oksydacyjne tłuszczu, lecz na wzrost jego ekskrecji z organizmu. Badania wykazały, że wzrost spożycia wapnia do ok. 1300 mg na dobę, głównie w postaci odtłuszczonych produktów mlecznych, wywołuje wzrost wydalania tłuszczów o około 8,2 g na dobę, przy diecie zawierającej około 30% tłuszczu. Mechanizm ten polega prawdopodobnie na tworzeniu się nierozpuszczalnych soli kwasów tłuszczowych z wapniem, które ograniczają absorpcję tłuszczu (50). Wykazano również, że wapń z przetworów mlecznych powoduje zmniejszenie poposiłkowej lipemii, prawdopodobnie na skutek redukcji absorpcji tłuszczu. Z kolei podawanie suplementów diety w postaci trudno rozpuszczalnych węglanów wapnia nie daje podobnych efektów (51). Powyższe założenia są zgodne z wynikami badań Zemela i wsp., którzy doszli do wniosku, że lepszy efekt daje wapń pochodzący z produktów nabiałowych niż wapń zawarty w suplementach diety zawierających węglany wapnia (49). Należy jednak wziąć pod uwagę, iż silniejszy wpływ jonów wapnia zawartych w produktach nabiałowych może wynikać z obecności w mleku i jego przetworach dodatkowych składników pokarmowych, których oddziaływanie z wapniem, może potęgować jego działanie (52).
Liczne badania dowodzą słuszności w spożywaniu większej ilości produktów nabiałowych w celu zapewnienia odpowiedniego poziomu wapnia, którego niedobór zwiększa ryzyko wystąpienia zespołu metabolicznego. Simin i wsp. zbadali grupę kobiet w wieku powyżej 45 lat i doszli do wniosku, że powszechność występowania zespołu metabolicznego była większa wśród pacjentek z niską podażą wapnia (pochodzącego zarówno z pożywienia, jak i suplementów diety) i odwrotnie (53).
Biorąc pod uwagę fakt, iż witamina D odpowiada za wewnątrzkomórkową homeostazę wapnia, oba te komponenty są metabolicznie ze sobą związane (53). Dlatego też, zbadano zależność między koncentracją witaminy D w surowicy krwi a ryzykiem wystąpienia jednego z elementów zespołu metabolicznego – otyłości.
Zależność między poziomem witaminy D a otyłością
Wiele publikacji donosi o odwrotnej zależności między poziomem witaminy D a otyłością. Zaobserwowano, że niedobór witaminy D występuję częściej u ludzi otyłych (45, 54, 55). Oprócz negatywnej korelacji między poziomem witaminy D a wskaźnikiem BMI (53), zarejestrowano również odwrotną zależność między surowiczym stężeniem witaminy D a występowaniem otyłości typu centralnego (otyłości brzusznej, czyli otyłości jabłkowej) (54). Można by zadać pytanie: czy deficyt witaminy D związany jest z niedostateczną ekspozycją na promienie słoneczne, czy wynika z samej otyłości? Wortsman i wsp. zaobserwowali, że po ekspozycji na promieniowanie ultrafioletowe, poziom witaminy D3 w surowicy krwi wzrósł zarówno u pacjentów otyłych, jak i u osób z grupy kontrolnej. Biorąc pod uwagę fakt, iż osoby z nadwagą mają większą powierzchnię ciała wystawioną na działanie promieni słonecznych, powinny one produkować więcej witaminy D. Tymczasem, zarejestrowana koncentracja witaminy D w surowicy krwi była u nich o około 57% mniejsza niż w grupie kontrolnej (55). Wykazano, że zawartość 7-dehydrocholesterolu u osób z obu grup jest podobna (56), w związku z czym, ilość powstającego w skórze kalcitriolu jest również zbliżona zarówno u pacjentów otyłych, jak i tych z prawidłową wagą ciała. Założono więc, że otyłość nie zakłóca produkcji witaminy D w skórze. Ponieważ jednak rozpuszczalna w tłuszczach witamina D magazynowana jest w tkance tłuszczowej, wysunięto hipotezę, że grubsza, niż u osób bez otyłości, podskórna tkanka tłuszczowa zatrzymuje witaminę D utrudniając jej transport do krwioobiegu. Konsekwencją jest zmniejszenie jej stężenia w surowicy krwi, w porównaniu z osobami normalnej wagi. Z kolei witamina D podawana doustnie wydaje się być bardziej biodostępna, prawdopodobnie dlatego, że zaraz po absorpcji trafia poprzez układ limfatyczny do krwioobiegu. Wydaje się zatem, że doustna podaż witaminy D pozwoli na uzupełnienie jej niedoborów u ludzi z otyłością (54).
Zależność między poziomem witaminy D a ryzykiem wystąpienia objawów zespołu metabolicznego wśród osób otyłych
Botella-Carretero i wsp. przebadali 73 otyłych pacjentów z niedoborem oraz bez deficytu witaminy D i stwierdzili, że niedostateczne stężenie witaminy D było obecne wśród 50,7% badanych, zaś objawy zespołu metabolicznego u 63%. Deficyt witaminy był bardziej powszechny u otyłych uczestników badań z syndromem zespołu metabolicznego w porównaniu z pacjentami bez żadnych objawów. Rozwój zaburzeń metabolicznych wśród pacjentów z otyłością może wynikać z zależności między poziomem witaminy D w surowicy krwi, a poziomami HDL i triglicerydów. Jak się okazało, stężenie witaminy D w surowicy krwi jest pozytywnie skorelowane z poziomem HDL, zaś negatywnie ze stężeniem triglicerydów (45). Witamina D zdaje się być potrzebna do utrzymania odpowiedniego stężenia apolipoproteiny A-1, białka będącego głównym składnikiem HDL. Dlatego u osób z wysokim poziomem witaminy D obserwowano również podwyższony poziom białka A-1 oraz HDL i odwrotnie – niedobór witaminy D prowadził do spadku stężenia apolipoproteiny A-1 i HDL (57). Z kolei u hemodializowanych pacjentów leczonych kalcitriolem poziom triglicerydów zmalał (58). Podobny efekt na stężenie triglicerydów we krwi miała wysoka podaż wapnia z diety (59). Z powyższych doniesień wynika, że niedobór witaminy D zwiększa ryzyko nadmiernego spadku poziomu HDL, sprzyja wzrostowi triglicerydów we krwi, dyslipidemii, cukrzycy i innych chorób mieszczących się pod pojęciem zespołu metabolicznego. Dlatego tak ważne jest, aby poziom witaminy D był uzupełniany, szczególnie przez osoby otyłe, u których gruba warstwa podskórnej tkanki tłuszczowej izoluje witaminę D od krwioobiegu czyniąc ją mniej dostępną, o czym trzeba pamiętać dyskutując profilaktykę zespołu metabolicznego.
Badania Botella-Carretero i wsp. zdają się być bardzo przekonujące, gdyż zostały przeprowadzone w oparciu o grupę pacjentów z tymi samymi wartościami BMI i obwodem talii, koncentrując się tym samym tylko na zależności między stężeniem witaminy D i powszechnością występowania objawów zespołu metabolicznego u osób otyłych (45). We wcześniejszych badaniach naukowych nie uwzględniono tych parametrów wskazując jednocześnie na odwrotną zależność między surowiczym stężeniem witaminy D a BMI (53) oraz otyłością wisceralną (54). Tym samym utrudnione było zbadanie zależności tylko między zespołem metabolicznym i witaminą D wśród osób otyłych, gdyż do dyskusji wchodziły jeszcze różne wartości BMI.
Zależność między poziomem witaminy D a ryzykiem wystąpienia nadciśnienia
Choroba ta jest główną przyczyną śmierci w krajach rozwiniętych. W samych tylko Stanach Zjednoczonych na nadciśnienie cierpi około 65 milionów osób (60). Liczne publikacje wskazują, że jednym z czynników ryzyka sprzyjających rozwojowi chorób układu krążenia jest niedostateczne spożycie pokarmów bogatych w witaminę D, jak również unikanie ekspozycji na promieniowanie UVB, które stanowi główne jej źródło dla organizmu ludzkiego.
Badania przeprowadzone w Stanach Zjednoczonych pozwoliły zauważyć, iż poziom 25OHD w surowicy krwi mniejszy niż 30 ng/ml (<75 nmol/l), dominował szczególnie wśród kobiet, osób starszych powyżej 60. roku życia, mniejszości etnicznych oraz osób z wyraźnymi czynnikami ryzyka, takimi jak: otyłość, nadciśnienie, cukrzyca, hipertriglicerydemia, czy hipercholesterolemia. Zaobserwowano, że stężenie witaminy D w surowicy krwi było negatywnie skorelowane z ryzykiem wystąpienia którejś z wyżej wymienionych chorób (61). Początkowo sądzono, że niskie poziomy kalcitriolu są warunkiem sine qua non w patogenezie zespołu metabolicznego. Z drugiej jednak strony spekulowano, że niedobór 1,25(OH)2D może pełnić jedynie rolę markera dla innych, nie znanych nam jeszcze czynników sprzyjających powstawaniu zaburzeń w ciśnieniu krwi, metabolizmie lipidów, glukozy, syntezy kalcitriolu i innych (62). Obecnie wiadomo, że wpływ witaminy D na ryzyko chorób układu krążenia może się wiązać z jej oddziaływaniem na układ renina-angiotensyna, i że witamina D pełni tu rolę negatywnego regulatora wewnątrzwydzielniczego, poprzez hamowanie ekspresji genu kodującego reninę (63). Witamina D znana była z roli pozytywnego regulatora genów znajdujących się pod jej kontrolą. Jak wspomniano heterodimer VDR-RXR (ryc. 1) wiążąc się z sekwencją VDRE w obrębie promotora genu docelowego, stymuluje ekspresję odpowiednich białek (19). System renina-angiotensyna poprzez szereg reakcji pośrednich prowadzi do utrzymania odpowiedniego ciśnienia krwi, przy czym nadmierny wzrost syntezy reniny sprzyja rozwojowi nadciśnienia. Badania in vitro przy użyciu komórek granularnych nerek, dowiodły roli kalcitriolu i jego analogów w supresji syntezy konwertora angiotensyny (ACE). Obecność sekwencji response elements dla VDR jest warunkiem wyhamowania ekspresji ACE i hypotensyjnych efektów witaminy D (61).
Li i wsp. z badań na szczurach wywnioskowali, że kalcitriol hamuje syntezę reniny w komórkach granularnych nerek, podczas gdy inaktywacja 1,25(OH)2D zwiększa ekspresję ACE. Zaobserwowano, że u myszy pozbawionych genu VDRE (knock-out) ekspresja reniny i angiotensyny II była kilkakrotnie większa, prowadząc do nadciśnienia, przerostu mięśnia sercowego i wzrostu pragnienia (63).
Badania populacyjne przeprowadzone na kobietach w różnym wieku potwierdziły negatywną zależność między podażą witaminy D a ciśnieniem krwi. Kobiety, które otrzymywały dzienną dawkę witaminy D w wysokości nie przekraczającej 400 IU miały nieco większe ciśnienie krwi niż pacjentki, których dzienna porcja witaminy wynosiła ponad 400 IU (64). Podobne wnioski wysunęli Krause i wsp., według których ekspozycja na promieniowanie UVB i związany z nią wzrost syntezy witaminy D w skórze był skorelowany ze spadkiem ciśnienia krwi (65).
Powyżej zamieściliśmy fragment artykułu, do którego możesz uzyskać pełny dostęp.
Mam kod dostępu
- Aby uzyskać płatny dostęp do pełnej treści powyższego artykułu albo wszystkich artykułów (w zależności od wybranej opcji), należy wprowadzić kod.
- Wprowadzając kod, akceptują Państwo treść Regulaminu oraz potwierdzają zapoznanie się z nim.
- Aby kupić kod proszę skorzystać z jednej z poniższych opcji.
Opcja #1
29 zł
Wybieram
- dostęp do tego artykułu
- dostęp na 7 dni
uzyskany kod musi być wprowadzony na stronie artykułu, do którego został wykupiony
Opcja #2
69 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 30 dni
- najpopularniejsza opcja
Opcja #3
129 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 90 dni
- oszczędzasz 78 zł
Piśmiennictwo
1. Tjellesen L, Hummer L, Christiansen C, Rodbro P. Serum concentration of vitamin D metabolites during treatment with vitamin D2 and D3 in normal premenopausal women. Bone Miner 1986; 1: 407-13. 2. Armas LA, Hollis BW, Heaney RP. Vitamin D2 is much less effective than vitamin D3 in humans. J Clin Endocrinol Metab 2004; 89(11): 5387-91. 3. Dusso AS, Brown AJ, Slatopolsky E. Vitamin D. Am J Physiol Renal Physiol 2005; 289: F8-F28. 4. Burgaz A, Ăkesson A, Öster A, Michaëlsson K, Wolk A. Associations of diet, supplement use and ultraviolet B radiation exposure with vitamin D status in Swedish women during winter. Am J Clin Natur 2007; 86: 1399-404. 5. Holick MF. Vitamin D: A millenium perspective. J Cell Biochem 2003; 88: 296-307. 6. Holick MF. Vitamin D: importance in the prevention of cancers, type 1 diabetes, heart disease, and osteoporosis. Am J Clin Nutr 2004; 79: 362-71. 7. Webb AR, Kline L, Holick MF. Influence of season and latitude on the cutaneous synthesis of vitamin D3: exposure to winter sunlight in Boston and Edmonton will not promote vitamin D3 synthesis in human skin. J Clin Endocrinol Metab 1988; 67: 373-8. 8. Webb AR, Engelsen O. Calculated ultraviolet exposure levels for a healthy vitamin D status. Photochem Photobiol 2006; 82: 1697-703. 9. Clemens TL, Adams JS, Henderson SL, Holick MF. Increased skin pigment reduces the capacity of skin to synthesise vitamin D3. Lancet 1982; 1: 74-6. 10. MacLaughlin J, Holick MF. Aging decreases the capacity of human skin to produce vitamin D3. J Clin Invest 1985; 76: 1536-8. 11. Webb AR. Who, what, where and when-influences on cutaneous vitamin D synthesis. Prog Biophys Mol Biol 2006; 92: 17-25. 12. Vieth R. Vitamin D supplementation, 25-hydroxy?vitamin D concentrations, and safety. Am J Clin Nutr 1999; 69: 842-56. 13. Calvo MS, Whiting SJ, Barton CN. Vitamin D fortification in the United States and Canada: current status and data needs. Am J Clin Nutr 2004; 80: 1710S-6S. 14. Outila TA, Mattila PH, Piironen VI, Lamberg-Allardt CJ. Bioavailability of vitamin D from wild edible mushrooms (Cantharellus tubaeformis) as measured with a human bioassay. Am J Clin Nutr 1999; 69: 95-8. 15. Gloth FM 3rd, Alam W, Hollis B. Vitamin D vs broad spectrum phototherapy in the treatment of seasonal affective disorder. J Nutr Health Aging 1999; 3: 5-7. 16. Vieth R. Vitamin D supplementation, 25-hydroxy?vitamin D concentrations, and safety. Am J Clin Nutr 1999; 69: 842-56. 17. Zittermann A. Vitamin D in preventive medicine: are we ignoring the evidence? Br J Nutr 2003; 89: 552-72. 18. Glerup H, Mikkelsen K, Poulsen L, Hass E, Overbeck S, Thomsen J, Charles P, Eriksen EF. Commonly recommended daily intake of vitamin D is not sufficient if sunlight exposure is limited. J Intern Med 2000; 247: 260-8. 19. Mathieu C, Gysemans C, Giulietti A, Bouillon R. Vitamin D and diabetes. Diabetologia 2005; 48; 1247-57. 20. Pascussi JM, Robert A, Nguyen M, Walrant-Debray O, Garabedian M, Martin P, Pineau T, Saric J, Navarro F, Maurel P, Vilarem MJ. Possible involvement of pregnane X receptor-enhanced CYP24 expression in drug-induced osteomalacia. J Clin Invest 2005; 115: 177-86. 21. Rochel N, Wurtz JM, Mitschler A, Klaholz B, Moras D. The crystal structure of the nuclear receptor for vitamin D bound to its natural ligand. Mol Cell 2000; 5: 173-9. 22. Zmuda JM, Cauley JA, Ferrell RE. Molecular epidemiology of vitamin D receptor gene variants. Epidemiol Rev 2000; 22: 203-17. 23. Holick MF. Sunlight and vitamin D for bone health and prevention of autoimmune diseases, cancers and cardiovascular disease. Am J Clin Nutr 2004; 80: 1678S-88S. 24. Norman A.W. Vitamin D: The calcium homeostasis steroid hormone. Academic Press. 1979. 25. Holick MF. Phylogenetic and evolutionary aspects of vitamin D from phytoplankton to humans. Vertebrate Endocrinology: Fundamentals and biochemical implications. Academic Press 1989; 3: 7-43. 26. Dawson-Hughes B, Heaney RP, Holick MF, Lips P, Meunier PJ, Vieth R. Estimates of optimal vitamin D status. Osteoporos Int 2005; 16: 713-6. 27. Heckman GA, Papaioannou A, Sebaldt RJ, Ioannidis G, Petrie A, Goldsmith C, Adachi JD. Effect of vitamin D on bone mineral density of elderly patients with osteoporosis responding poorly to bisphosphonates. BMC Musculoskelet Disord 2002; 3: 6. 28. McAlindon TE, Felson DT, Zhang Y, Hannan MT, Aliabadi P, Weissman B, Rush D, Wilson PW, Jacques P. Relation of dietary intake and serum levels of vitamin D to progression of osteoarthritis of the knee among participants in the Framingham Study. Ann Intern Med 1996; 125: 353-9. 29. Glerup H, Mikkelsen K, Poulsen L, Hass E, Overbeck S, Andersen H, Charles P, Eriksen EF. Hypovitaminosis D myopathy without biochemical signs of osteomalacic bone involvement. Calcif Tissue Int 2000; 66: 419-24. 30. Arai H, Miyamoto K-I, Taketani Y. A vitamin D receptor gene polymorphism in the translation initiation codon: effect on protein activity and relation to bone mineral density in Japanese women. I Bone Miner Res 1997; 12: 915-21. 31. Geusens P, Vandevyver C, Vanhoof J, Cassiman JJ, Boonen S, Raus J. Quadriceps and grip strength are related to vitamin D receptor genotype in elderly nonobese women. I Bone Miner Res 1997; 12: 2082-8. 32. Hopkinson NS, Li KW, Kehoe A, Humphries SE, Roughton M, Moxham J, Montgomery H, Polkey MI. Vitamin D receptor genotypes influence quadriceps strength in chronic obstructive pulmonary disease. Am J Clin Nutr 2008; 87: 385-90. 33. Grundberg E, Brändström H, Ribom EL, Ljunggren O, Mallmin H, Kindmark A. Genetic variation in the human vitamin D receptor is associated with muscle strength, fat mass and body weight in Swedish women. Eur J Endocrinol 2004; 150: 323-8. 34. Holick MF. Noncalcemic actions of 1,25-dihydroxyvitamin D3 and clinical applications. Bone 1995; 17: 107S-11S. 35. Endo I, Inoue D, Mitsui T, Umaki Y, Akaike M, Yoshizawa T, Kato S, Matsumoto T. Deletion of vitamin D receptor gene in mice results in abnormal skeletal muscle development with deregulated expression of myoregulatory transcription factors. Endocrinology 2003; 144: 5138-44. 36. Bischoff-Ferrari HA, Borchers M, Gudat F, Durmuller U, Stahelin HB, Dick W. Vitamin D receptor expression in human muscle tissue decreases with age. J Bone Miner Res 2004; 19: 265-9. 37. Garland CF, Garland FC, Gorham ED, Lipkin M, Newmark H, Mohr SB, Holick MF. The role of vitamin D in cancer prevention. Am J Public Health 2006; 96: 252-61. 38. Smedby KE, Hjalgrim H, Melbye M, Torrang A, Rostgaard K, Munksgaard L, Adami J, Hansen M, Porwit-MacDonald A, Jensen BA, Roos G, Pedersen BB, Sundström C, Glimelius B, Adami HO. Ultraviolet radiation exposure and risk of malignant lymphomas. J Natl Cancer Inst 2005; 97: 199-209. 39. Nagpal S, Na S, Rathnachalam R. Noncalcemic actions of vitamin D receptor ligands. Endocr Rev 2005; 26: 662-87. 40. Naveilhan P, Berger F, Haddad K, Barbot N, Benabid AL, Brachet P, Wion D. Induction of glioma cell death by 1,25(OH)2 vitamin D3: towards an endocrine therapy of brain tumors? J Neurosci Res 1994; 37: 271-7. 41. Trouillas P, Honnorat J, Bret P, Jouvet A, Gerard JP. Redifferentiation therapy in brain tumors: long-lasting complete regression of glioblastomas and an anaplastic astrocytoma under long term 1-alpha-hydroxycholecalciferol. J Neurooncol 2001; 51: 57-66. 42. Woo TC, Choo R, Jamieson M, Chander S, Vieth R. Pilot study: potential role of vitamin D (Cholecalciferol) in patients with PSA relapse after definitive therapy. Nutr Cancer 2005; 51: 32-6. 43. Mantell DJ, Owens PE, Bundred NJ, Mawer EB, Canfield AE. 1á25-dihydroxyvitamin D3 inhibits angiogenesis in vitro and in vivo. Circ Res 2000; 87: 214-20. 44. Peters U, McGlynn KA, Chatterjee N, Gunter E, Garcia-Closas M, Rothman N, Sinha R. Vitamin D, calcium, and vitamin D receptor polymorphism in colorectal adenomas. Cancer Epidemiol Biomarkers Prev 2001; 10: 1267-74. 45. Botella-Carretero JI, Alvarez-Blasco F, Villafruela JJ, Balsa JA, Vázquez C, Escobar-Morreale HF. Vitamin D deficiency is associated with the metabolic syndrome in morbid obesity. Clin Nutr 2007; 26: 573-80. 46. Liu S, Song Y, Ford ES, Manson JE, Buring JE, Ridker PM. Dietary calcium, vitamin D and the prevalence of metabolic syndrome In middle-aged and older U.S. women. Diabetes Care 2005; 28: 2926-32. 47. Chiu KC, Chu A, Go VL, Saad MF. Hypovitaminosis D is associated with insulin resistance and â cell dysfunction. Am J Clin Nutr 2004; 79: 820-5. 48. Davies KM, Heaney RP, Recker RR, Lappe JM, Barger-Lux MJ, Rafferty K, Hinders S. Calcium intake and body weight. J Clin Endocrinol Metab 2000; 85: 4635-8. 49. Zemel MB. Regulation of adiposity and obesity risk by dietary calcium: mechanisms and implications. J Am Coll Nutr 2002; 21: 146S-51S. 50. Jacobsen R, Lorenzen JK, Toubro S, Krog-Mikkelsen I, Astrup A. Effect of short-term high dietary calcium intake on 24-h energy expenditure, fat oxidation, and fecal fat excretion. Int J Obes (Lond) 2005; 29: 292-301. 51. Lorenzen JK, Nielsen S, Holst JJ, Tetens I, Rehfeld JF, Astrup A. Effect of dairy calcium or supplementary calcium intake on postprandial fat metabolism, appetite, and subsequent energy intake. Am J Clin Nutr 2007; 85: 678-87. 52. Davies KM, Heaney RP, Recker RR, Lappe JM, Barger-Lux MJ, Rafferty K, Hinders S. Calcium intake and body weight. J Clin Endocrinol Metab 2000; 85: 4635-8. 53. Liu S, Song Y, Ford ES, Manson JE, Buring JE, Ridker PM. Dietary calcium, vitamin D and the prevalence of metabolic syndrome In middle-aged and older U.S. women. Diabetes Care 2005; 28: 2926-32. 54. Ford ES. Ajani UA, McGuire LC, Liu S. Concentrations of serum vitamin D and the metabolic syndrome among U.S. adults. Diabetes Care 2005; 28: 1228-30. 55. Wortsman J, Matsuoka LY, Chen TC, Lu Z, Holick MF. Decreased bioavailability of vitamin D in obesity. Am J Clin Nutr 2000; 72: 690-3. 56. Need AG, Morris HA, Horowitz M, Nordin C. Effects of skin thickness, age, body fat, and sunlight on serum 25-hydroxyvitamin D. Am J Clin Nutr 1993; 58: 882-5. 57. Auwerx J, Bouillon R, Kesteloot H. Relation between 25-hydroxyvitamin D3, apolipoprotein A-1 and high density lipoprotein cholesterol. Arterioscler Thromb 1992; 12: 671-4. 58. Lin SH, Lin SYF, Lu KC. Effects of intravenous calcitriol on lipid profiles and glucose tolerance in uraemic patients with secondary hyperparathyroidism. Clin Sci (Lond) 1994; 87: 533-8. 59. Papakonstantinou E, Flaut WP, Huth PJ, Harris RB. High dietary calcium reduces body fat content, digestibility of fat, and serum vitamin D in rats. Obes Res 2003; 11: 387-94. 60. Forman JP, Bischoff-Ferrari HA, Willett WC, Stampfer MJ, Curhan GC. Vitamin D intake and risk of incident hypertension results from three large prospective cohort studies. Hypertension 2005; 46: 676-82. 61. Martins D, Wolf M, Pan D, Zadshir A, Tareen N, Thadhani R, Felsenfeld A, Levine B, Mehrotra R, Norris K. Prevalence of cardiovascular risk factors and the serum levels of 25-hydroxyvitamin D in the United States. Arch Intern Med 2007; 167: 1159-65. 62. Lind L, Hänni A, Lithell H, Hvarfner A, Sörensen OH, Ljunghall S. Vitamin D is related to blood pressure and other cardiovascular risk factors in middle-aged men. Am J Hypertens 1995; 8: 894-901. 63. Li YC, Kong J, Wei M, Chen ZF, Liu SQ, Cao LP. 1,25-dihydroxyvitamin D3 is a negative endocrine regulator of the renin-angiotensin system. J Clin Invest 2002; 110: 229-38. 64. Sowers MR, Wallace RB, Lemke JH. The association of intakes of vitamin and calcium with blood pressure among women. Am J Clin Nutr 1985; 42: 135-42. 65. Krause R, Bühring M, Hopfenmüller W, Holick MF, Sharma AM. Ultraviolet B and blood pressure. Lancet 1998; 352: 709-10. 66. Pfeifer M, Begerow B, Minne HW, Nachtigall D, Hansen C. Effects of a short-term vitamin D3 and calcium supplementation on blood pressure and parathyroid hormone levels in elderly women. J Clin Endocrinol Metab 2001; 86: 1633-7. 67. McCarron DA, Reusser ME. Finding Consensus in the dietary calcium-blood pressure debate. J Am Coll Nutr 1999; 18: 398S-405S. 68. Pittas AG, Lau J, Hu FB, Dawson-Hughes B. The role of vitamin D and calcium in type 2 diabetes. A systematic review and meta-analysis. J Clin Endocrinol Metab 2007; 92: 2017-29. 69. Johnson JA, Grande JP, Roche PC, Kumar R. Immunohistochemical localization of the 1,25(OH)2D3 receptor and calbindin D28k in human and rat pancreas. Am J Physiol 1994; 267: E356-E60. 70. Zeitz U, Weber K, Soegiarto DW, Wolf E, Balling R, Erben RG. Impaired insulin secretory capacity in mice lacking a functional vitamin D receptor. FASEB J 2003; 17: 509-11. 71. Bland R, Markovic D, Hills CE, Hughes SV, Chan SL, Squires PE, Hewison M. Expression of 25-hydroxyvitamin D3-1alpha-hydroxylase in pancreatic islets. J Steroid Biochem Mol Biol 2004; 89-90: 121-5. 72. Maestro B, Dávila N, Carranza MC, Calle C. Identification of a vitamin D response element in the human insulin receptor gene promoter. J Steroid Biochem Mol Biol 2003; 84: 223-30. 73. Bland R, Markovic D, Hills CE, Hughes SV, Chan SL, Squires PE, Hewison M. Expression of 25-hydroxyvitamin D3-1-hydroxylase in pancreatic islets. J Steroid Biochem Mol Biol 2004; 89-90: 121-5. 74. Norman AW, Frankel JB, Heldt AM, Grodsky GM. Vitamin D deficiency inhibits pancreatic secretion of insulin. Science 1980; 209: 823-5. 75. Cade C, Norman AW. Vitamin D3 improves impaired glucose tolerance and insulin secretion in the vitamin D-deficient rat in vivo. Endocrinology 1986; 119: 84-90. 76. Clark SA, Stumpf WE, Sar M. Effect of 1,25-dihydroxyvitamin D3 on insulin secretion. Diabetes 1981; 30: 382-6. 77. Chiu KC, Chu A, Go VL, Saad MF. Hypovitaminosis D is associated with insulin resistance and â cell dysfunction. Am J Clin Nutr 2004; 79: 820-5. 78. Sooy K, Schermerhorn T, Noda M, Surana M, Rhoten WB, Meyer M, Fleischer N, Sharp GW, Christakos S. Calbindin-D(28k) controls [Ca(2+)](i) and insulin release. Evidence obtained from calbindin-d(28k) knockout mice and beta cell lines. J Biol Chem 1999; 274: 34343-9. 79. The Eurodiab Substudy 2 Study Group. Vitamin D supplement in early childhood and risk for type I (insulin-dependent) diabetes mellitus. Diabetologia 1999; 42: 51-4. 80. Stene LC, Joner G, Norwegian Childhood Diabetes Study Group. Use of cod liver oil during the first year of life is associated with lower risk of childhood-onset type 1 diabetes: a large, population-based, case-control study. Am J Clin Nutr 2003; 78: 1128-34. 81. Hyppönen E, Läärä E, Reunanen A, Järvelin MR, Virtanen SM. Intake of vitamin D and risk of type 1 diabetes: a birth-cohort study. Lancet 2001; 358: 1500-3. 82. Cantorna MT, Zhu Y, Froicu M, Wittke A. Vitamin D status, 1,25-dihydroxyvitamin D3 and the immune system. Am J Clin Nutr 2004; 80: 1717S-20S. 83. Hahn HJ, Kuttler B, Mathieu C, Bouillon R. 1,25-dihydroxyvitamin D3 reduces MHC antigen expression on pancreatic beta-cells in vitro. Transplant Proc 1997; 29: 2156-7. 84. Towers TL, Staeva TP, Freedman LP. A two-hit mechanism for vitamin D3-mediated transcriptional repression of the granulocyte-macrophage colony-stimulating factor gene: vitamin D receptor competes for DNA binding with NFAT1 and stabilizes c-Jun. Mol Cell Biol 1999; 19: 4191-9. 85. Gregori S, Giarratana N, Smiroldo S, Uskokovic M, Adorini L. A 1alpha,25-dihydroxyvitamin D(3) analog enhances regulatory T-cells and arrests autoimmune diabetes in NOD mice. Diabetes 2002; 51: 1367-74. 86. Casteels KM, Gysemans CA, Waer M, Bouillon R, Laureys JM, Depovere J, Mathieu C. Sex difference in resistance to dexamethasone-induced apoptosis in NOD mice: treatment with 1,25(OH)2D3 restores defect. Diabetes 1998; 47: 1033-7. 87. Mathieu C, Waer M, Casteels K, Laureys J, Bouillon R. Prevention of type I diabetes in NOD mice by nonhypercalcemic doses of a new structural analog of 1,25-dihydroxyvitamin D3, KH1060. Endocrinology 1995; 136: 866-72. 88. Munger KL, Zhang SM, O´Reilly E, Hernán MA, Olek MJ, Willett WC, Ascherio A. Vitamin D intake and incidence of multiple sclerosis. Neurology 2004; 62: 60-5. 89. Richards JB, Valdes AM, Gardner JP, Paximadas D, Kimura M, Nessa A, Lu X, Surdulescu GL, Swaminathan R, Spector TD, Aviv A. Higher serum vitamin D concentrations are associated with longer leukocyte telomere length in women. Am J Clin Nutr 2007; 86: 1420-5. 90. Oelzner P, Muller A, Deschner F, Huller M, Abendroth K, Hein G, Stein G. Relationship between disease activity and serum levels of vitamin D metabolites and PTH in rheumatoid arthritis. Calcif Tissue Int 1998; 62: 193-8. 91. Richards JB, Valdes AM, Gardner JP, Paximadas D, Kimura M, Nessa A, Lu X, Surdulescu GL, Swaminathan R, Spector TD, Aviv A. Higher serum vitamin D concentrations are associated with longer leukocyte telomere length in women. Am J Clin Nutr 2007; 86: 1420-5. 92. Kurz DJ, Decary S, Hong Y, Trivier E, Akhmedov A, Erusalimsky JD. Chronic oxidative stress compromises telomere integrity and accelerates the onset of senescence in human endothelial cells. J Cell Sci 2004; 117: 2417-26. 93. van der Harst P, van der Steege G, de Boer RA, Voors AA, Hall AS, Mulder MJ, van Gilst WH, van Veldhuisen DJ; MERIT-HF Study Group. Telomere length of circulating leukocytes is decreased in patients with chronic heart failure. J Am Coll Cardiol 2007; 49: 1459-64. 94. Valdes AM, Andrew T, Gardner JP, Kimura M, Oelsner E, Cherkas LF, Aviv A, Spector TD. Obesity, cigarette smoking, and telomere length in women. Lancet 2005; 366: 662-4. 95. Laaksi I, Ruohola JP, Tuohimaa P, Auvinen A, Haataja R, Pihlajamaki H, Ylikomi T. An association of serum vitamin D concentrations < 40 nmol/L with acute respiratory tract infection in young Finnish men. Am J Clin Nutr 2007; 86: 714-7. 96. Liu PT, Stenger S, Li H, Wenzel L, Tan BH, Krutzik SR, Ochoa MT, Schauber J, Wu K, Meinken C, Kamen DL, Wagner M, Bals R, Steinmeyer A, Zugel U, Gallo RL, Eisenberg D, Hewison M, Hollis BW, Adams JS, Bloom BR, Modlin RL. Toll-like receptor triggering of a vitamin D-mediated human antimicrobial response. Science 2006; 311: 1770-3. 97. Bischoff-Ferrari HA, Dietrich T, Orav EJ, Dawson-Hughes B. Positive association between 25-hydroxy vitamin D levels and bone mineral density: a population-based study of younger and older adults. Am J Med 2004; 116: 634-9. 98. Johansson C, Black D, Johnell O, Odén A, Mellström D. Bone mineral density is a predictor of survival. Calcif Tissue Int 1998; 63: 190-6. 99. Visser M, Deeg DJ, Puts MT, Seidell JC, Lips P. Low serum concentrations of 25-hydroxyvitamin D in older persons and the risk of nursing home admission. Am J Clin Nutr 2006; 84: 616-22. 100. Snijder MB, van Dam RM, Visser M, Deeg DJ, Dekker JM, Bouter LM, Seidell JC, Lips P. Adiposity in relation to vitamin D status and parathyroid hormone levels: a population-based study in older men and women. J Clin Endocrinol Metab 2005; 90: 4119-23. 101. Lind L, Wengle B, Ljunghall S. Blood pressure is lowered by vitamin D (alphacalcidol) during long-term treatment of patients with intermittent hypercalcaemia. A double-blind, placebo-controlled study. Acta Med Scand 1987; 222: 423-7.
