© Borgis - Medycyna Rodzinna 2/2008, s. 48-60
Justyna Kosińska1, Katarzyna Billing-Marczak1, *Marcin Krotkiewski2
Nowopoznana rola witaminy K w patogenezie chorób cywilizacyjnych
Newly discovered role of vitamin k in the pathogenesis of diseases of civilization
1Dział Badań i Rozwoju Nowych Produktów, Biotech-Varsovia Sp. z o.o. – Sp. K. w Warszawie Dyrektor Działu: dr n. biol. Katarzyna Billing-Marczak 2The Sahlgrenska Academy at Göteborg University, Institute of Clinical Neuroscience, Göteborg, Sweden
Summary
Vitamins belonging to K family comprise of several compounds showing similar structure divided into K1 (Phylloquinone), K2 (Menaquinones) and K3 (Menadione). Apart from well known function of vitamin K as important coagulation factor, vitamin K shows a high spectrum of activities even in extrahepatic tissues.
Vitamin K takes part in carboxylation of proteins participating in mineralization of bones and calcification of blood vessels. There are osteocalcin produced by osteoblasts and MGP (Matrix Gla Protein) produced by epithelial cells in blood vessels and by chondrocytes in cartilage. Osteocalcin increases mineralization of bones, MGP prevents calcification of blood vessels. Moreover, vitamin K stimulates synthesis of collagen proteins. Vitamin K increases production of osteoblasts and apoptosis of osteoclasts changing the balance between these two types of cells towards formation of new bone tissue. Epidemiological studies showed negative correlation between the development of osteoporosis and cardiovascular diseases and the plasma concentration of vitamin K. Several studies indicate that vitamin K acts effectively in the prophylaxis and treatment of osteoporosis and arteriosclerosis.
Vitamin K inhibits polymerase γ activity in mitochondria and thereby induces the oxidative stress and apoptosis in cancer cells. It causes also the increase of expression of protooncogenes and TR3 protein as well as the inhibition of connexin CX43 depressing the proliferation and inducing massive apoptosis of tumor cells. In addition vitamin K plays an important role in the limitation of impairment of neuron functions and decline of cognitive brain functions after stroke. It modulates synthesis of sphingolipids and is proposed to be used in the prophylaxis of Alzeimer´s disease.

WSTĘP
Odkrycia witaminy K w roku 1930 dokonał duński biochemik Henrik Dam. Analizując metabolizm cholesterolu u kurczaków zauważył on, iż utrzymywane na niskotłuszczowej i pozbawionej steroli diecie kurczaki wykazywały rozległe krwawienia w tkance podskórnej i w mięśniach (1, 2). Ponadto, McFarlane i wsp. zauważyli, że zwierzęta karmione mięsem ryb lub mięsem wieprzowym, z którego wyekstrahowano tłuszcze, cierpiały na zaburzenia krzepliwości krwi nawet po krótkim pobycie w laboratorium (3). Rozpoczęto intensywne badania różnych typów pasz i produktów żywnościowych (4). W 1934 roku Dam i Schönheyder włączając lub wykluczając z diety kurczaków poszczególne komponenty zaobserwowali, że dieta bogata w zielone liście, pomidory oraz wątrobę wieprzową zapobiegała krwawieniom. Wysunięto hipotezę, iż zaburzenia w krzepliwości krwi są wynikiem niedoboru czynnika rozpuszczalnego w tłuszczach (2), którego głównym źródłem jest wątroba wieprzowa. Henryk Dam nazwał ów czynnik witaminą koagulacji (w skrócie witaminą K) biorąc pod uwagę jej antykrwotoczny charakter (4).
W drugiej połowie lat 30. szereg grup badawczych podjęło się prób izolacji witaminy K z różnego rodzaju produktów żywnościowych. Jak się okazało, bogatym jej źródłem jest lucerna (alfalfa), z której wyekstrahowano witaminę K w postaci żółtego oleju (2, 5, 6) oraz mięso ryb, z którego naukowcy z St. Louis pod kierunkiem Edwarda Doisy wyizolowali czynnik antykrwotoczny w postaci krystalicznej (7). W 1935 roku Almquist i Stokstad odkryli, że witamina K pochodząca z wyżej wymienionych źródeł powstaje w wyniku działania bakterii jelitowych, takich jak Escherichia coli. Potwierdziły to badania nad zespołem krwawień u kurczaków, które wykazały, że podawanie ekstraktu z lucerny, mięsa ryb lub ekstraktu z owsa poddane fermentacji bakteryjnej mogą działać profilaktycznie i zapobiegać krwawieniom (8). Ponadto, Dam i wsp. wykazali, że wyekstrahowany z zielonych liści preparat olejowy, podawany noworodkom ze skłonnością do krwawień wynikającą z niskiego poziomu protrombiny, wykazywał działanie profilaktyczne zmniejszając śmiertelność okołoporodową z 4,6% do 1,8% (2). Z biegiem czasu podawanie witaminy K noworodkom i ich matkom stało się coraz bardziej powszechne (9). O odkryciu prewencyjnej roli witaminy K w zespole krwawień u noworodków, Dam wspomniał podczas wręczania nagrody Nobla, którą otrzymał wraz z Doisy w 1943 za odkrycie witaminy K i ustalenie jej struktury chemicznej (2).
BUDOWA WITAMINY K
Niedługo po odkryciu czynnika koagulacji ustalono jego strukturę chemiczną oraz zidentyfikowano dwie naturalne jego formy. Pierwszą z nich wyizolowaną z zielonych liści lucerny nazwano witaminą K1, drugą – wyekstrahowaną z rybiego mięsa – witaminą K2 (2). Obecnie wiemy, że witamina K2 to nazwa dla całej grupy czynników koagulacyjnych, która obejmuje szereg związków wykazujących niewielkie różnice w budowie molekularnej.
Cechą charakterystyczną witamin z grupy K jest wspólny dla wszystkich pierścień 2-metylo-1,4-naftochinonowy z przyłączonymi w pozycji C-3 grupami izoprenoidowymi. Te ostatnie stanowią podstawę klasyfikacji witamin K do różnych rodzin (K1, K2 i K3) w zależności od ich liczby (ryc. 1) (10).
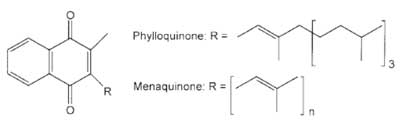
Ryc. 1. Struktura filochinonu (witamina K1) i menachinonów (witaminy K2) (10).
Witamina K1, czyli filochinon (ang. Phylloquinone) lub fitomenadion (ang. Phytonadione) zawiera jedną nienasyconą i trzy nasycone grupy izoprenoidowe. Witamina K1 syntetyzowana jest wyłącznie w roślinach.
Witamina K2 to cała grupa związków określanych mianem menachinonów (ang. Menaquinones). Ich struktura została ustalona w latach 60. XX wieku (11), a nomenklatura bazuje na ilości grup izoprenoidowych w łańcuchu bocznym. Menachinony (zwane w skrócie MK-n, gdzie n oznacza liczbę nienasyconych grup izoprenoidowych w pozycji 3 i waha się w granicach od n = 1 do n = 13) z większą liczbą tych grup (np. MK-7, czy MK-10), syntetyzowane są przez bakterie i występują głównie w produktach pochodzenia zwierzęcego (10, 12), a w Japonii – także w produktach fermentacji soi, takich jak natto. MK-4 jest jedynym menachinonem produkowanym w komórkach ssaków w wyniku konwersji witaminy K1, MK-7 (11) oraz witaminy K3 (10, 13). Badania wykazały, że doustne podawanie witaminy K1 pozwala na jej przekształcenie w witaminę K2 poprzez usunięcie alifatycznego łańcucha bocznego z cząsteczki filochinonu. Co ciekawe, konwersja K1 do K2 nie zachodzi w przypadku podaży witaminy K1 drogą dojelitową lub do płynu mózgowo-rdzeniowego (14).
Witamina K3, czyli menadion (ang. Menadione) jest syntetycznym, pozbawionym alifatycznego łańcucha bocznego, analogiem witaminy K pełniącym rolę prowitaminy. Menadion charakteryzuje wysoka aktywność biologiczna i zdolność do konwersji w MK-4 w komórkach ssaków (12, 13).
ŹRÓDŁA WITAMINY K
Liczne badania wykazały, że najczęściej spotykaną w żywności witaminą K jest filochinon (15), który w diecie zachodniej stanowi ponad 80% całkowitej podaży witaminy K (16). Pozostały procent to witaminy z grupy K2, których źródłem jest nie tylko dieta, ale również inne postaci witamin K ulegające konwersji do MK-4. Dlatego też uważa się, że zarówno filochinon, jak i menachinony w równym stopniu mogą zasilać pulę witaminy K w ludzkim organizmie (10).
Syntetyzowana wyłącznie w roślinach (10), witamina K1 występuje głównie w zielonych liściach różnych roślin, zielonych jarzynach (11), olejach roślinnych i margarynach (13), zaś najmniejszą ilość filochinonu stwierdzono w mięsie i produktach nabiałowych. Zawartość witaminy K1 waha się w szerokim zakresie i zależy od źródła, w którym występuje: zielone warzywa (100-750 ?g/100 g), margaryna i oleje roślinne (50-200 ?g/100 g) oraz Natto (20-40 ?g/100 g) (15).
Menachinony syntetyzowane przez bakterie, występują głównie w produktach pochodzenia zwierzęcego, takich jak mięso, jaja i produkty mleczne (10, 15). Szczególnie bogate w menachinony są także produkty fermentacji soi, takie jak Natto, gdzie dominującą formą witaminy jest MK-7, syntetyzowany z udziałem Bacillus subtilis (13, 15). Najważniejsze menachinony w diecie obejmują grupy od MK-4 do MK-10 (11). Zawartość MK-4 w mięsie waha się w granicach 1-30 ?g/100 g, w jajach 10-25 ?g/100 g, w innych produktach nabiałowych 0,2-15 ?g/100 g, zaś koncentracja pozostałych menachinonów (MK-n) sięga 900-1200 ?g na 100 g Natto, 40-80 ?g na 100 g serów oraz do 35 ?g na 100 g innych produktów nabiałowych (15).
PODAŻ WITAMINY K
Rekomendowane dzienne spożycie (RDA) witaminy K wynosi 1 ?g/1 kg masy ciała. Dawka ta została oparta na ilościach witaminy potrzebnych do syntezy czynników biorących udział w procesach krzepnięcia krwi. Jednak wybitną zdolnością do klirensu witaminy K, przy małych jej ilościach, charakteryzuje się tylko wątroba, podczas gdy akumulacja witaminy K w innych organach (serce, trzustka, nerki, płuca, aorta) następuje dopiero przy wyższej jej podaży (17). Dlatego też obecnie rekomendowane dzienne spożycie witaminy K jest wystarczające jedynie do utrzymania funkcji krzepnięcia krwi, zaś zupełnie niewystarczające do pokrycia potrzeb w innych tkankach.
Liczne badania wskazują na to, że rekomendowane dzienne spożycie witaminy K zależy od rodzaju tkanki i organu, jak również parametrów takich jak przepływ krwi i gęstość receptorów apolipoproteiny (apoE) na powierzchni komórek docelowych. Ilości docierających do różnych organów witamin z grupy K zależą bowiem między innymi od nośników, z pomocą których są transportowane. I tak, witamina K1 związana z frakcją trójglicerydów przenoszona jest głównie do wątroby, zaś nośnikiem transportującym witaminy K2 do innych tkanek są frakcje LDL i HDL lipoprotein (10). Badania wskazują, że o ile witamina K1 (naturalna) wychwytywana jest przez wątrobę dość szybko (zaledwie podczas pierwszych 8 godzin po jej podaniu), tak MK-7 (pochodzący z Natto) podlega najpierw redystrybucji wątrobowej, po której transportowany w krwiobiegu z pomocą lipoprotein LDL i HDL staje się dostępny dla tkanek pozawątrobowych. Potwierdzają to badania nad czasem połowicznego rozpadu, według których okres półtrwania witaminy K1 jest znacznie krótszy w porównaniu do MK-7 i wynosi zaledwie 1,5 godziny. Koncentracja menachinonu w krwiobiegu była z kolei możliwa do zarejestrowania nawet po 72 godzinach po jego podaniu. Okres półtrwania syntetycznej witaminy K1 jest nieco dłuższy od jej naturalnego analogu, jednak krótszy w porównaniu z MK-7 pochodzącym z Natto. Uważa się, że różnica w sposobie transportu, jak i okresie półtrwania wspomnianych witamin sprawia, że MK-7 zdaje się być bardziej i dłużej dostępny dla tkanek pozawątrobowych, takich jak tkanka kostna i ściany naczyń krwionośnych niż filochinon, czy MK-4 (16). Schurgers i wsp. w swojej pracy przeglądowej donoszą, że u szczurów efekty menachinonów o dłuższych łańcuchach bocznych, jak MK-9, dzięki ich dłuższemu okresowi półtrwania, utrzymywały się przez znacznie dłuższy okres czasu niż efekty działania filochinonu (15).
W związku z powyższym, wielkość dawki jaka powinna być stosowana dla poszczególnych form witaminy K, powinna być dostosowana z uwzględnieniem ich różnych czasów połowicznego rozpadu. Badania wskazują, że standaryzowana dawka MK-4, przy której efekty kliniczne są porównywalne z tymi, jakie obserwowano po podaniu Natto, wynosi 45 mg/dzień. Stanowi to około 1000-krotnie większą dawkę w porównaniu z 45 ?g MK-7 na dzień. Wynika to prawdopodobnie z faktu, że menachinony o dłuższych łańcuchach bocznych wykazują dłuższy okres półtrwania (mniejsze tempo klirensu wątrobowego) i pozostają dłużej dostępne dla innych tkanek (efektywniejsze wiązanie z lipoproteinami przez dłuższy okres czasu) niż menachinony, takie jak MK-4 (16). Naukowcy zakładają, że w przypadku witaminy K1 właściwą dawką powinna być ilość na poziomie około 1 mg/dzień (18, 19). Badania porównawcze przeprowadzone dla MK-7 i witaminy K1 dowodzą, że przy tych samych dawkach obu witamin poziom filochinonu w surowicy krwi był około 6-7 razy mniejszy niż MK-7. Potwierdziły to analizy, według których koncentracja nieukarboksylowanej osteokalcyny (ucOC) była mniejsza przy podaży MK-7 niż filochinonu, zaś stosunek ukarboksylowanej do nieukarboksylowanej osteokalcyny (cOC/ucOC) wykazywał tendencję wzrostową nawet po 6. tygodniach stosowania MK-7. Zależność cOC/ucOC przy podaży witaminy K1 wzrastała jedynie do 2. tygodni podczas jej podawania (16, 20). W związku z powyższym słusznym wydaje się stwierdzenie, że zalecana dobowa dawka witaminy K powinna być dostosowana do poszczególnych jej form z uwzględnieniem wyższych dawek dla filochinonu i menachinonów o krótszych łańcuchach bocznych. Jednocześnie, bardzo niska toksyczność i pozytywne efekty jakie witamina K wywiera na układ kostny i naczynia krwionośne (21) zdają się być racjonalną podstawą do ponownego rozpatrzenia rekomendowanych dawek witaminy, jak i ewentualnego wzbogacania w nią produktów i dodatków żywieniowych.
Należy również podkreślić, iż wchłanianie jelitowe nie jest identyczne dla obu witamin. Podczas gdy absorpcja filochinonu sięga zaledwie 5-15% całkowitej przyjętej z pokarmem witaminy K1, tak witamina K2 wchłaniana jest niemal w całości (10). Koncentrując się na biodostępności witaminy K przeprowadzono doświadczenie, z którego wywnioskowano, że doustna podaż tych samych ilości syntetycznej witaminy K1 i naturalnego MK-7 pochodzącego z Natto prowadzi ostatecznie do niemalże 10-krotnie większego surowiczego stężenia MK-7 względem filochinonu (16). Różnica ta prawdopodobnie wynika z rodzaju źródła, z którego pochodzi dana postać witaminy. Występujący w roślinach filochinon, ściśle związany z membranami chloroplastów, znacznie trudniej ulega absorpcji, niż menachinony wchłanianie głównie z tłuszczów i produktów nabiałowych (15).
Podaż witaminy K a szerokość geograficzna. Podaż witaminy K nie jest jednakowa na całym świecie. Badania przeprowadzone w Ameryce Północnej, Europie i Azji wykazały, że rozbieżność w spożyciu witaminy K jest skorelowana z szerokością geograficzną, różnymi populacjami oraz wiekiem. W Irlandii 1/3 kobiet w okresie pomenopauzalnym nie przyjmuje rekomendowanych ilości filochinonu, który stanowi podstawową formę witaminy K zawartą w diecie w krajach Ameryki Północnej i Europy (22). Badania przeprowadzone na ludności holenderskiej powyżej 55. roku życia wykazały, że szacunkowe spożycie witaminy K1 waha się w granicach od 124 ?g do 375 ?g na dzień, zaś witaminy K2 między 10 ?g a 45 ?g na dzień (10). Wysoka podaż witaminy K w diecie Holendrów może wynikać z wysokiej konsumpcji warzyw przez tę populację (15). Podobnie, spożycie witaminy K1 wśród osób starszych zamieszkujących Chiny było znacznie wyższe w porównaniu z ludnością Wielkiej Brytanii, co może potwierdzać związek między większą podażą witaminy K a zwiększoną konsumpcją warzyw (23) i produktów fermentacji soi na Dalekim Wschodzie.
ORGANY DOCELOWE DLA WITAMINY K
Głównym magazynem dla witamin z grupy K jest wątroba, do której transportowana jest większość wchłoniętej z pokarmu witaminy. Pozostałe tkanki gromadzą witaminę K dopiero przy wyższej jej podaży (17). Ponadto, stosunek koncentracji filochinonu do menachinonów wykazuje duże zróżnicowanie w zależności od rodzaju tkanki (10). I tak, zawartość witaminy K1 w wątrobie jest około 10-krotnie wyższa niż MK-4 (17). Organem z równie wysoką koncentracją filochinonu jest mięsień serca, podczas gdy wysoką akumulacją witaminy K2 charakteryzują się trzustka, jądra oraz ściany tętnic. Badania wykazały, że w tkankach tych koncentracja menachinonów utrzymuje się na wysokim poziomie nawet przy długoterminowej suplementacji samą witaminą K1. Hipoteza, że filochinon ulega konwersji do MK-4 z udziałem flory bakteryjnej została poddana w wątpliwość po przeprowadzeniu badań, na podstawie których stwierdzono, że stężenie MK-4 u wyjałowionych szczurów było utrzymywane na wysokim poziomie mimo podaży samej witaminy K1. Zakłada się, że 90% wchłoniętego filochinonu ulega konwersji do MK-4 w przypadku, gdy podaż menachinonów w diecie jest po prostu niewystarczająca (10). Badania przeprowadzone z udziałem osób starszych sugerują, że magazynem dla filochinonu i menachinonów może być również tkanka kostna (24).
CYKL WITAMINY K (OKSYDACJA -? REDUKCJA)
Witamina K jest niezbędnym czynnikiem zaangażowanym w posttranslacyjną modyfikację białek. Proces ten polega na γ-karboksylacji reszt glutaminowych (Glu) w efekcie czego (10) powstają reszty kwasu γ-karboksyglutaminowego (Gla), wykazujące powinowactwo do jonów wapnia. Dzięki tym ostatnim, dojrzałe już białka uzyskują zdolność wiązania jonów wapnia, co czyni je funkcjonalnie aktywnymi. Zmodyfikowane tą drogą białka noszą nazwę białek Gla (10, 25, 26). γ-karboksylacja zachodzi przy udziale systemu integralnych białek błon retikulum endoplazmatycznego (ER), w skład którego wchodzą enzymy zaangażowane w cykl przemian witaminy K oraz konwersję kwasu glutaminowego (Glu) w kwas γ-karboksyglutaminowy (Gla) w obrębie modyfikowanych białek (10, 26). Do enzymów tych należą: zależna od witaminy K γ-karboksylaza oraz wrażliwa na warfarynę reduktaza epoksydowa (ang. Vitamin K 2,3-epoxide reductase; VKOR). Pierwszy enzym wymaga obecności zredukowanej formy witaminy K (KH2), drugi – odpowiada za konwersję witaminy K do postaci zredukowanej (KH2). Cykl przemian witaminy K oraz proces karboksylacji białek Gla są nierozerwalnie ze sobą powiązane. Każda cząsteczka przechodzi przez następujące po sobie etapy oksydacji i redukcji (10). Formą aktywną witaminy K jest postać zredukowana (KH2), która pełni rolę kofaktora dla γ-karboksylazy zaangażowanej w proces konwersji reszt Glu do reszt Gla (13). Następująca jednocześnie oksydacja KH2 (ang. Vitamin K – hydroquinone) do formy utlenionej KO (ang. Vitamin K – epoxide) dostarcza energii potrzebnej do tego procesu (27). W kolejnym etapie KO ulega redukcji do KH2 z udziałem tioli (12) i reduktazy VKOR zamykając cykl przemian witaminy K (26, 27). Tym samym zostaje odtworzona postać aktywna witaminy K, która może ponownie uczestniczyć w kolejnym cyklu. Proces recyklingu witaminy K sprawia, że każda cząsteczka może być wykorzystana nawet kilka tysięcy razy, co tłumaczy jej niewielkie dzienne zapotrzebowanie w porównaniu z innymi witaminami (ryc. 2) (10).
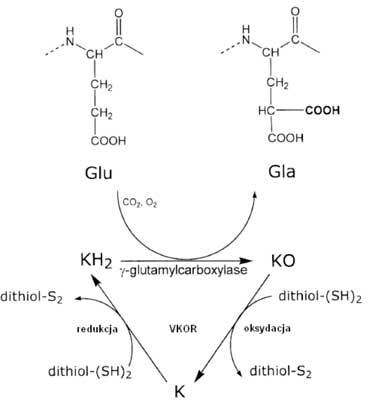
Ryc. 2. Cykl witaminy K. Glu – kwas glutaminowy; Gla – kwas γ-karboksyglutaminowy; KH2 – postać zredukowana, aktywna witaminy K; KO – postać utleniona witaminy K; VKOR – reduktaza epoksydowa (10).
ROLA BIAŁEK GLA
Istnienie specyficznego aminokwasu jakim jest kwas γ-karboksyglutaminowy (Gla) po raz pierwszy stwierdzono w połowie lat 70. XX wieku. Białkiem, w którym znaleziono Gla była protrombina (11). Obecność aminokwasu Gla w protrombinie jest warunkiem niezbędnym do zajścia zależnej od wapnia konwersji tego białka do postaci aktywnej – trombiny (28). Z biegiem czasu, aminokwas ten zidentyfikowano również w innych czynnikach biorących udział w procesie krzepnięcia krwi (11). Obecnie wiemy, że witamina K jest konieczna do posttranslacyjnej modyfikacji białek syntetyzowanych w wątrobie, takich jak: protrombina, czynniki krzepnięcia krwi (VII, IX, X) oraz białka S, C i Z (2, 10, 28, 29). Co ciekawe, obecność zależnej od witaminy K karboksylazy kwasu glutaminowego odkryto także w skórze, kościach (30) i chrząstkach, co wskazuje, że endogenne substraty dla tego enzymu to nie tylko białka odpowiedzialne za krzepliwość krwi (11). Obecnie wiadomo o istnieniu przynajmniej 16 białek zawierających reszty Gla u ssaków, z czego 8 zaangażowanych jest w proces krzepnięcia krwi, zaś rola pozostałych wykracza poza układ krwionośny (30). Zakłada się przy tym, że antagoniści witaminy K, np. kumaryny, mogą negatywnie wpływać na aktywność karboksylazy kwasu glutaminowego także w tkankach pozawątrobowych poprzez blokadę reduktazy epoksydowej i zaburzenie cyklu regeneracji witaminy K (11). Suttie w swojej pracy przeglądowej donosi o obecności enzymu karboksylazy również w innych organach, takich jak śledziona, trzustka, płuca, łożysko, jądra, tarczyca, grasica, ścięgna, macica, nerki, a także w fibroblastach i komórkach nowotworowych różnego pochodzenia (28). Z powyższych wynika, że występowanie białek Gla obejmuje szeroki zakres tkanek i organów w całym organizmie (31), z czym wiąże się również ich zróżnicowanie funkcjonalne. Zależne od witaminy K białka uczestniczą między innymi w metabolizmie kości (21, 32, 33), metabolizmie naczyń krwionośnych, prewencji zwapnienia naczyń (13, 32) i kalcyfikacji tkanek miękkich, w procesach wzrostu i apoptozy komórek (33) oraz adhezji komórkowej (13, 32).
Do najlepiej zbadanych białek Gla należą osteokalcyna i MGP (ang. Matrix Gla Protein) (10), których rola została poznana w oparciu o badania z udziałem myszy transgenicznych. Dowiedziono, że zwierzęta pozbawione genów dla osteoklacyny lub MGP wykazują zaburzenia w mechanizmach kalcyfikacji wielu tkanek. Jak się później okazało, różnica między nimi polega na tkankach docelowych, na jakie każde z tych białek oddziałuje: osteokalcyna zorientowana jest na układ kostny (34), zaś MGP – na tkankę chrzęstną i ściany naczyń krwionośnych (31, 35). Funkcje białek Gla skorelowane są z miejscem ich syntezy, która ma miejsce w osteoblastach (10, 31) i odontoblastach (36) – w przypadku osteokalcyny lub w komórkach tkanki chrzęstnej (chondrocytach), mięśniówce gładkiej naczyń (10, 31) i w śródbłonku naczyń – w przypadku MGP.
Funkcja witaminy K jako kofaktora dla enzymu odpowiedzialnego za posttranslacyjną modyfikację białek obejmuje również szereg białek będących ligandami dla receptorów białkowych kinaz tyrozynowych (ang. Receptor Tyrosine Kinases; RTK) (12). Rola ligandów w aktywacji wspomnianych receptorów obejmuje udział w procesach komórkowych, takich jak: wzrost i przeżywalność komórek, transformacja komórkowa, czy migracja komórek. Funkcję tę dzielą dwa zależne od witaminy K białka: Gas6 (ang. Growth-arrest-specific-6) i białko S (37).
LECZENIE PRZECIWZAKRZEPOWE
Doustne środki przeciwzakrzepowe prowadzą do wystąpienia wielu efektów ubocznych. W Polsce najbardziej znane antykoagulanty to warfaryna i acenokumarol. Związki te, łącząc się z reduktazą epoksydową VKOR, hamują recykling witaminy K na etapie redukcji KO do KH2 w rezultacie czego, karboksylacja wielu zależnych od witaminy K białek zostaje zahamowana. Nieukarboksylowane białka Gla, zwane PIVKA (ang. Proteins Induced by Vitamin K Absence and Antagonists), pozostają funkcjonalnie nieaktywne prowadząc do zaburzeń obserwowanych w różnych tkankach i organach. Badania przeprowadzone na większej populacji ujawniły, że wysokie poziomy niedokarboksylowanej osteokalcyny są skorelowane z niedomineralizowaniem kości i osteoporozą (38), zaś wysoka koncentracja niedokarboksylowanego białka MPG – z wysokim stopniem zwapnienia tętnic (39).
Istnienie powikłań w terapii warfarynowej zwiększa znaczenie alternatywnych metod leczenia przeciwzakrzepowego poprzez podawanie pacjentom leków takich jak: aspiryna, klopidogrel, dipyramidol, czy też inhibitory trombiny i czynnika X (11). Dyskutowano również jednoczesne podawanie witaminy K i warfaryny. Jednakże w przypadku tego typu terapii istnieje potencjalne niebezpieczeństwo związane z faktem, że witamina K1 wychwytywana jest głównie przez wątrobę, podczas gdy inne tkanki i organy są bardziej podatne na jej niedobór. Zatem zwiększona dawka warfaryny podawana w celu skompensowania zwiększonej podaży witaminy K może prowadzić do niedoborów witaminy K w ścianach naczyń krwionośnych, zwiększając ryzyko zwapnienia naczyń. Hipotezę tę potwierdziły badania na zwierzętach, według których szczury otrzymujące witaminę K1 w połączeniu z warfaryną wykazywały intensywne zwapnienie naczyń (40). Naukowcy sugerują, że w celu uniknięcia efektu zwapnienia naczyń rozwiązaniem wydaje się być podawanie witaminy K2, która nie wychwytywana przez wątrobę w takich ilościach jak filochinon, transportowana jest również do innych organów w ilościach odpowiadających jej podaży. Potwierdziły to badania, według których podawanie szczurom MK-4 i warfaryny nie prowadziło do kalcyfikacji naczyń (41). Inni naukowcy spekulują, iż celem uniknięcia powikłań w terapii przeciwzakrzepowej optymalne wydaje się być podawanie MK-7 ze względu na dłuższy okres połowicznego rozpadu w porównaniu do filochinonu (42).
ROLA WITAMINY K W METABOLIZMIE KOŚCI
Jedne z pierwszych danych o zaangażowaniu witaminy K w metabolizm tkanki kostnej zanotowano w latach 70. XX wieku. Obserwacje te dotyczyły deformacji układu szkieletowego u dzieci matek leczonych antykoagulantami, takimi jak warfaryna, w czasie pierwszego trymestru ciąży (43). Jednocześnie, Pastoureau i wsp. zaobserwowali, że otrzymujące warfarynę owce już po kilku miesiącach wykazywały wyraźne cechy osteopenii (44), zaś analiza wyników badań u pacjentów poddanych długoterminowej terapii przeciwzakrzepowej ujawniła znaczne zmniejszenie masy kostnej i słabsze uwapnienie układu szkieletowego (45).
Obecnie wiemy, że metabolizm tkanki kostnej jest ściśle związany z prawidłowym poziomem witaminy K, której rola nie ogranicza się jedynie do procesów krzepnięcia krwi, lecz obejmuje również procesy przebudowy kości. Liczne badania wskazują na udział witaminy K2 w tłumieniu resorpcji kości poprzez hamowanie aktywności i indukowanie apoptozy osteoklastów przy jednoczesnej inhibicji apoptozy osteoblastów (46) i indukcji różnicowania komórek progenitorowych w komórki kościotwórcze (47). W efekcie takiego działania, witamina K2 przesuwa równowagę w kierunku formowania tkanki kostnej, a nie jej ubytku.
Zaangażowanie witaminy K w utrzymanie właściwej homeostazy tkanki kostnej odzwierciedlają zależne od posttranslacyjnej karboksylacji białka. Najbardziej reprezentatywnym białkiem układu kostnego po kolagenie jest osteokalcyna. Białko to, syntetyzowane przez osteoblasty, należy do białek niekolagenowych wchodzących w skład ECM (ang. Extracellular Matrix) (48). Osteokalcyna posiada trzy reszty Gla i odgrywa istotną rolę w wiązaniu jonów wapnia do hydroksyapatytu (36) – nieorganicznego składnika kości, który stanowi około 70% całkowitej masy kostnej. W postaci nieukarboksylowanej osteoklacyna pozostaje nieaktywna i niezdolna do wbudowywania minerałów w tkankę kostną, w rezultacie czego dochodzi do zmniejszenia masy kostnej i zwiększenia podatności na złamania (16, 49). Poziom nieukarboksylowanej osteokalcyny odzwierciedla koncentrację witaminy K (36). Jednocześnie stężenie w surowicy krwi nieukarboksylowanego białka wzrasta wraz z wiekiem i jest wyraźnie skorelowane z ryzykiem złamań kości (50). Z tego też powodu, osteokalcyna uważana jest za marker aktywności osteoblastów (11). Z każdym rokiem przybywa danych epidemiologicznych wskazujących na zależność między niedoborem witaminy K, poziomem nieukarboksylowanej osteokalcyny oraz związaną z wiekiem utratę masy kostnej i zwiększone ryzyko złamań kości na tle osteoporozy. Szulc i wsp. potwierdzili istnienie zależności między wysokim poziomem nieukarboksylowanej osteokalcyny a zwiększonym ryzykiem złamań kości (38, 51) oraz utratą masy kostnej u starszych kobiet (52). Feskanish i wsp. w oparciu o wyniki dziesięcioletniego badania donoszą, że kobiety w wieku między 38. a 63. rokiem życia otrzymujące najniższe dawki witaminy K (poniżej 109 ?g/dzień) charakteryzowały się znacznie większym ryzykiem złamania kości biodrowej (53). Jednocześnie, przeprowadzone w Danii badania na kobietach w wieku pomenopauzalnym wykazały, że suplementacja filochinonu w ilości 200 ?g/dzień i 500 ?g/dzień prowadziła po 6 tygodniach do redukcji koncentracji nieukarboksylowanej osteokalcyny o odpowiednio 40% i 68% (22). Osteokalcyna jest białkiem zależnym od witaminy K, zatem wysoki poziom nieukarboksylowanego białka wskazuje na niedobory witaminy K i spowodowane tym zwiększone ryzyko złamań kości. Liczne badania podkreślają, że wysokie spożycie witaminy K jest niezależnym czynnikiem obniżającym ryzyko wystąpienia osteoporozy i złamań kości w okresie pomenopauzalnym.
Wiadomo również, że witamina K uczestniczy w modyfikacji posttranslacyjnej szeregu innych białek, m.in uczestniczącej w pozakomórkowej mineralizacji kości periostyny (54).
Jednakże z innych badań wynika, że fenotypowe odchylenia od normy w układzie kostnym myszy pozbawionych osteokalcyny i MGP nie do końca potwierdzają ogólnie przyjęty pogląd o istnieniu zależności protekcyjnych funkcji witaminy K na układ kostny od modyfikacji białek szkieletowych. Badania bowiem wskazują, iż brak ekspresji MGP lub osteokalcyny u myszy również prowadził do wzrostu masy kostnej, a nie jej ubytku. Wyniki te dowodzą, iż rola witaminy K w metabolizmie tkanki kostnej zachodzi również inną drogą, nie tylko poprzez karboksylację białek Gla (55). Jak się okazało, witamina K2 odpowiada również za wzrost produkcji specyficznej dla układu kostnego fosfatazy alkalicznej (ALP) oraz innych markerów wzmożonej aktywności osteoblastów, takich jak: MGP (ang. Matrix Gla Protein), OPG (ang. Osteoprotegerin; Osteoclastogenesis-inhibitory Factor) i OPN (ang. Osteopontin). Tabb i wsp. w oparciu o przeprowadzone badania wysunęli hipotezę, że regulacja transkrypcji markerów tkanki kostnej zachodząca za pośrednictwem witaminy K2 związana jest z aktywacją receptora SXR (46). SXR (ang. Steroid and Xenobiotic Receptor) i jego homolog PXR u myszy (ang. Pregnane X Receptor) (55) jest jądrowym receptorem aktywującym transkrypcję specyficznych genów w odpowiedzi na stymulację przez szereg substancji biologicznych i ksenobiotycznych. Ekspresja SXR ma miejsce m.in. w układzie szkieletowym, gdzie odgrywa istotną rolę w procesach przebudowy tkanki kostnej (36). Witamina K2 będąc bardzo silnym agonistą SXR, poprzez stymulację receptora, moduluje ekspresję wielu genów kodujących ważne dla układu kostnego białka. Grupa Horie-Inoue przedstawiła nowy mechanizm działania witaminy K w układzie kostnym, według którego witamina K2 reguluje transkrypcję genów kodujących białka matrycy pozakomórkowej, tzw. ECM (ang. Extracellular Matrix) w zależny od SXR sposób oraz zwiększa akumulację kolagenu w osteoblastach (56). W systemie hodowli ludzkich osteoblastów linii MG63, która wykazuje nieustanną ekspresję SXR, zidentyfikowano spokrewnione z ECM geny tsukushi (TSK) i matrilin-2 (MATN2). Tsukushi to gen należący do rodziny proteoglikanów SLRR (ang. Small Leucine-Rich Repeat) (57), która odpowiada za wzrost syntezy włókien kolagenowych (58). Matryliny, uczestniczą w tworzeniu elementów włóknistych zależnych od produkcji kolagenu (59). Zakłada się, że witamina K oddziałując na SXR promuje syntezę kolagenu w osteoblastach poprzez regulację transkrypcji genów kodujących białka macierzy pozakomórkowej, TSK i MATN2 (36). Sugeruje się więc, że witamina K2 wpływa na poprawę jakości tkanki kostnej poprzez oddziaływanie na drogę sygnałową SXR (ryc. 3).
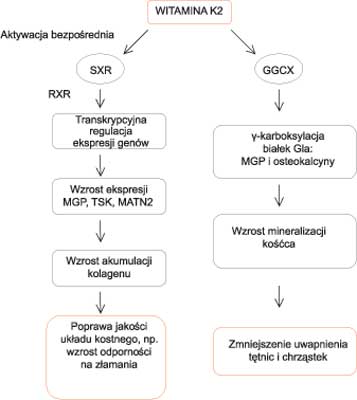
Ryc. 3. Podwójna rola witaminy K.
1) Receptor SXR po utworzeniu heteodimeru z RXR i aktywowaniu witaminą K2, promuje akumulację kolagenu w osteoblastach poprzez regulację transkrypcji genów kodujących białka matrycy zewnątrzkomórkowej.
2) Witamina K2 uczestniczy w posttranslacyjnej modyfikacji białek Gla pełniąc rolę kofaktora dla enzymu γ-glutamylkarboksylazy (GGCX). Aktywne białka Gla odpowiadają za wzrost mineralizacji kośćca i zmniejszenie uwapnienia naczyń krwionośnych (36).
Obecnie rozważaną kwestią jest włączenie agonistów receptora SXR do repertuaru terapeutycznego chorób układu kostnego, takich jak osteoporoza (36).
Jednocześnie zaobserwowano, iż receptor SXR nie jest jedynym czynnikiem niezbędnym do ekspresji osteoprotekcyjnych efektów witaminy K. Istotną rolę odgrywa również Msx2 będący osteoblastycznym czynnikiem transkrypcyjnym zidentyfikowanym jako gen indukowany menachinonem 4. Igarashi i wsp. wykazali, iż osteoblastogeneza indukowana w komórkach linii MC3T3-E1 przez MK-4 jest zależna od PXR i Msx2. Wyciszenie genu Msx2 z pomocą siRNA prowadziło do zniesienia efektów działania witaminy K2. Podobny rezultat uzyskano po wyłączeniu genu dla PXR, co znacznie osłabiło różnicowanie komórek prekursorowych w osteoblasty. Zidentyfikowano PXRE (ang. PXR Response Elements) w promotorze genu Msx2, co sugeruje jego zależność od PXR, a tym samym witaminy K. Igarashi i wsp. sugerują, że Msx2 jest genem docelowym dla receptora PXR aktywowanego witaminą K. Z powyższych wynika, że witamina K działa na poziomie genomu poprzez aktywację genu Msx2 odpowiedzialnego za różnicowanie osteoblastów z komórek prekursorowych (55).
Interesujące wnioski pochodzą z badania Framingham Offspring Study, z którego wynika negatywna korelacja między poziomem witaminy K i stężeniem markerów stanu zapalnego, takich jak IL-6, osteoprotegryna i TNF-α. Stan zapalny towarzyszący schorzeniom układu krążenia i osteoporozie wyraża się wzrostem syntezy prozapalnych cytokin. Badania nad zależnością między koncentracją filochinonu w surowicy krwi oraz jego podażą a stężeniem markerów stanu zapalnego wykazały odwrotną korelację między nimi sugerując możliwy udział witaminy K w reakcjach zapalnych organizmu. Jednocześnie brak zależności między procentem nieukarboksylowanej osteokalcyny a poziomem czynników prozapalnych pozwala przypuszczać, że witamina K moduluje stan zapalny w sposób niezależny od jej roli w procesie karboksylacji zależnych od niej białek. Spekuluje się, że witamina K hamuje stan zapalny poprzez bezpośrednie oddziaływanie i zmniejszenie ekspresji genów poszczególnych cytokin, takich jak IL-6 i osteoprotegryna. Podobną, odwrotną zależność zaobserwowano w przypadku suplementacji witaminy D, która prowadziła do zmniejszenia stężenia markerów stanu zapalnego. Z powyższych wynika przypuszczalna ochronna rola witamin K i D w łagodzeniu stanów zapalnych towarzyszących schorzeniom m.in. układu krążenia i tkanki kostnej (60).
ROLA WITAMINY K W PREWENCJI ZWAPNIENIA NACZYŃ I INNE EFEKTY POZA UKŁADEM KRZEPNIĘCIA
Zwapnienie naczyń jest istotnym czynnikiem prowadzącym do rozwoju chorób układu krążenia, takich jak miażdżyca na skutek redukcji elastyczności ścian naczyń krwionośnych, a tym samym wzrostu ich sztywności i ciśnienia tętniczego. U ludzi odkładanie soli wapniowo-fosforanowych w tkankach sercowo-naczyniowych (33, 61) zachodzi w dwojaki sposób. Magazynowanie wapnia w najbardziej wewnętrznej warstwie ścian naczyń krwionośnych, tzw. tunica intima, jest postacią zwapnienia najczęściej powiązanego ze zmianami miażdżycowymi, podczas gdy kalcyfikacja środkowej warstwy ścian naczyń, tzw. tunica media (zwana również sklerozą Mönckeberga) może występować nawet u osób bez zaawansowanych zmian miażdżycowych. Ta postać zwapnienia często towarzyszy schorzeniom, takim jak ciężka niewydolność nerek i cukrzyca (10).
Początkowo uważano, że proces wapnienia naczyń polega na biernym odkładaniu wapnia w postaci hydroksyapatytu (podobnie jak w tkance kostnej) w miejscach degradacji komórkowej. Obecnie wiadomo, że kalcyfikacja naczyń jest procesem aktywnym (32) związanym z charakterystyczną cechą komórek mięśni gładkich naczyń, tzw. VSMC (ang. Vascular Smooth Muscle Cell), które mogą nabywać zdolność do ekspresji białek charakterystycznych dla osteoblastów (10) i chondrocytów (32). Zważywszy na fakt, iż w ścianach wapniejących naczyń zidentyfikowano wiele białek typowych dla tkanek kostnej i chrzęstnej (osteokalcyna, MGP, osteopontyna, osteonektyna, białka morfogenetyczne kości) wydaje się słuszne stwierdzenie, że kalcyfikacja naczyń jest procesem przypominającym uwapnienie kości (10).
Powyżej zamieściliśmy fragment artykułu, do którego możesz uzyskać pełny dostęp.
Mam kod dostępu
- Aby uzyskać płatny dostęp do pełnej treści powyższego artykułu albo wszystkich artykułów (w zależności od wybranej opcji), należy wprowadzić kod.
- Wprowadzając kod, akceptują Państwo treść Regulaminu oraz potwierdzają zapoznanie się z nim.
- Aby kupić kod proszę skorzystać z jednej z poniższych opcji.
Opcja #1
29 zł
Wybieram
- dostęp do tego artykułu
- dostęp na 7 dni
uzyskany kod musi być wprowadzony na stronie artykułu, do którego został wykupiony
Opcja #2
69 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 30 dni
- najpopularniejsza opcja
Opcja #3
129 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 90 dni
- oszczędzasz 78 zł
Piśmiennictwo
1. Dam H, Schönheyder F: A deficiency disease in chicks resembling scurvy. Biochem J 1934; 28: 1355-9. 2. Zetterstrom R. H. C. P. Dam (1895-1976), E. A. Doisy (1893-1986): The discovery of antihaemorrhagic vitamin and its impact on neonatal health. Acta Paediatrica 2006; 95: 642-4. 3. McFarlane WD, Graham WR, Richardson F: The FAT-soluble vitamin requirements of the chick: the vitamin A and vitamin D content of fish meal and meat meal. Biochem J 1930; 25: 358-66. 4. Dam H: The antihaemorrhagic vitamin of the chick. Biochem J 1935; 29: 1273-85. 5. Almquist HJ: Purification of the antihemorrhagic vitamin. J Biol Chem 1936; 114: 241-5. 6. Almquist HJ: Purification of the antihemorrhagic vitamin by distillation. J Biol Chem 1936; 115: 589-91. 7. McKee RW, Binkley SB, MacCorquodale DW: The isolation of vitamins K1 and K2. J Am Chem Soc 1936; 61: 1295. 8. Almquist HJ, Stokstad ELR: Hemorrhagic chick disease of dietary origin. J Biol Chem 1935; 111: 105-13. 9. Lehmann J: Vitamin K as a prophylactic In 13.000 infants. Lancet 1944; 243: 493-4. 10. Vermeer C, Braam L: Role of K vitamins in the regulation of tissue calcification. J Bone Miner Metab 2001; 19: 201-6. 11. Cranenburg ECM, Schurgers LJ Vermeer C: Vitamin K: The coagulation vitamin that became omnipotent. Thromb Haemost 2007; 98: 120-5. 12. Lamson DW, Plaza SM: The anticancer effects of vitamin K. Alternative Medicine Review 2003; 8: 303-18. 13. Vermeer C et al.: Beyond Deficiency: Potential benefits of increased intakes of vitamin K for bone and vascular health. Eur J Nutr 2004; 43: 325-35. 14. Okano T et al.: Conversion of Phylloquinone (Vitamin K1) into Menaquinone-4 (Vitamin K2) in Mice. Two possible routes for menaquinone+4 accumulation in cerebra of mice. J Biol Chem 2008; 283: 11270-9. 15. Schurgers LJ et al.: Nutritional intake of vitamin K1 (Phylloquinone) and K2 (Menaquinone) in The Netherlands. Journal of Nutritional & Enviromental Medicine 1999; 9: 115-22. 16. Opracowanie: Benefits Beyond coagulation. Effects on osteoporosis and calcification of arteries. NAttoPharma. 17. Ronden JE, Thijssen HWH, Vermeer C: Tissue distribution of K-vitamers under different nutritional regimens in the rat. Biochim Biophys Acta Acta 1998; 1379: 16-22. 18. Knapen MH, Hamulyak K, Vermeer C: The effect of vitamin K supplementation on circulating osteocalcin (bone Gla protein) and urinary calcium excretion. Ann Int Med 1989; 111: 1001-5. 19. Schurgers LJ, Vermeer C: Differential lipoprotein transport pathways of K-vitamins in healthy subjects. Biochim Biophys Acta 2002; 1570: 27-32. 20. Vermeer C: Serum level of absorbed vitamin MK-7 and K1 in healthy people aged 55-60 years. Pilot Study 2; 2003. 21. Adams J, Pepping J: Vitamin K In the treatment and prevention of osteoporosis and arterial calcification. Am J Health-Syst Pharm 2005; 62: 1574-81. 22. Collins A, Cashman KD, Kiely M: Phylloquinone (vitamin K1) intakes and serum undercarboxylated osteocalcin levels In Irish postmenopausal women. Br J Nutr 2006; 95: 982-8. 23. Booth SL: Vitamin K status In the elderly. Curr Opin Clin Nutr Metab Care 2007; 10: 20-9. 24. Hodges SJ et al.: Detection and measurements of vitamins K1 and K2 in human cortical and trabecular bone. J Bone Miner Res 1993; 8: 1005-8. 25. Proudfoot D, Shanahan CM: Molecular mechanism mediating vascular calcification: Role of matrix Gla protein. Nephrology 2006; 11: 455-61. 26. Wallin R, Sane DC, Hutson SM: Vitamin K 2,3-epoxide reductase and the vitamin K-dependent g-carboxylation system. Thromb Res 2003; 108: 221-6. 27. Schurgers LJ et al.: Regression of warfarin-induced medial elastocalcinosis by high intake of vitamin K in rats. Blood 2007; 109: 2823-31. 28. Suttie JW: Vitamin K-dependent carboxylase. Ann Rev 1985; 54: 459-77. 29. Cain D, Hutson SM, Wallin R: Assembly of the warfarin-sensitive vitamin K 2,3-epoxide reductase enzyme complex in the endoplasmic reticulum membrane. J Biol Chem 1997;272: 29068-75. 30. Chu PH et al.: Purified vitamin K epoxide reductase alone is sufficient for conversion of vitamin K epoxide to vitamin K and vitamin K to vitamin KH2. PNAS 2006; 103: 19308-13. 31. Cranenburg ECM et al.: The circulating inactive form of Matrix Gla Protein (ucMGP) as a biomarker for cardiovascular Calcification. J Vasc Res 2008; 45: 427-36. 32. Shearer MJ: Role of vitamin K and Gla proteins in the pathophysiology of osteoporosis and vascular calcification. Curr Opin Clin Nutr & Metab Care 2000; 3: 433-8. 33. Erkkila AT, Booth SL: Vitamin K intake and atherosclerosis. Curr Opin Lipidol 2008; 19: 39-42. 34. Karsenty G: Genetics of Skeletogenesis. Dev Genet 1998; 22: 301-13. 35. Luo G et al.: Spontaneous calcification of arteries and cartilage In mice lacking matrix Gla protein. Nature 1997; 386: 78-81. 36. Horie-Inoue K, Inoue S: Steroid and xenobiotic receptor mediates a novel vitamin K2 signaling pathway in osteoblastic cells. J Bone Miner Metab 2008; 26: 9-12. 37. Saxena SP, Israels ED, Israels LG: Novel vitamin K dependent pathways regulating cell survival. Apoptosis 2001; 6: 57-68. 38. Szulc P et al.: Serum undercarboxylated osteocalcin is a marker of the risk of hip frapture: a three year follow-up study. Bone 1996; 18: 487-8. 39. Braam LA et al.: Assay for human Matrix Gla Protein in serum: potential application in the cardiovascular field. Arterioscler Thromb Vasc Biol 2000; 20: 1257-61. 40. Price PA, Faus SA, Williamson MK: Warfarin causes rapid calcification of the elastic lamellae in rat arteries and heart valves. Arterioscler Thromb Vasc Biol 1998; 18: 1400-7. 41. Spronk HM et al.: Tissue-specific utilization of menaquinone-4 results in the prevention of arterial calcification in warfarin-treated rats. J Vasc Res 2003; 40: 531-7. 42. Stafford DW, Roberts HR, Vermeer C: Vitamin K supplementation during oral anticoagulation: cautions. Blood 2007; 109: 3607. 43. Lietman PS, Pettifor JM, Benson R: Congenital malformations associated with the administration of oral anticoagulants during pregnancy. J Pediatr 1975; 86: 459-62. 44. Pastoureau P et al.: Osteopenia and bone-remodeling abnormalities in warfarin-treated lambs. J Bone Miner Res 1993; 8: 1417-26. 45. Gage BF et al.: Risk of osteoporotic fracture in elderly patients taking warfarin: results from the National Registry of Atrial Fibrillation 2. Arch Intern Med 2006; 166: 241-6. 46. Tabb MM, Sun A, Zhou C: Vitamin K2 Regulation of Bone Homeostasis Is Mediated by the Steroid and Xenobiotic Receptor SXR. J Biol Chem 2003; 278: 43919-27. 47. Koshihara Y et al.: Vitamin K stimulates osteoblastogenesis and inhibits osteoclastogenesis in human bone marrow cell culture. J Endocrinol 2003; 3: 339-48. 48. Ducy P et al.: Increased bone formation In osteocalcin-deficient mice. Nature 1996; 382: 448-52. 49. Booth SL et al.: Effects of vitamin K supplementation on bone loss In elderly men and women. J Clin Endocrinol Metab 2008; 93: 1217-23. 50. Vergnaud P et al.: Undercarboxylated osteocalcin measured with a specific immunoassay predicts hip fracture in elderly women: the EPIDOS Study. J Clin Endocrinol Metab 1997; 82: 719-24. 51. Szulc P et al.: Serum undercarboxylated osteocalcin is a marker of the risk of hip fracture in elderly women. J Clin Invest 1993; 4: 1769-74. 52. Szulc P et al.: Serum undercarboxylated osteocalcin correlates with hip bone mineral density in elderly women. J Bone Miner Res 1994; 9: 1591-5. 53. Feskanish D et al.: Vitamin K intake and hip fracture In women: a prospective study. Am J Clin Nutr 1999; 1: 74-9. 54. Coutu D et al.: Periostin: a member of a novel family of vitamin K-dependent proteins is expressed by mesenchymal stromal cells. J Biol Chem 2008; 1-21. 55. Igarashi M et al.: Vitamin K induces osteoblast differentiation through pregnane X receptor-mediated transcriptional control of the Msx2 gene. Mol Cell Biol 2007; 27: 7947-54. 56. Ichikawa T et al.: Steroid and Xenobiotic Receptor SXR Mediates Vitamin K2-activated Transcription of Extracellular Matrix-related Genes and Collagen Accumulation in Osteoblastic Cells. J Biol Chem 2006; 281: 16927-34. 57. Ohta K et al.: Tsukushi functions as an organizer inducer by inhibition of BMP activity in cooperation with chordin. Dev Cell 2004; 7: 347-58. 58. Wiberg C et al.: Complexes of Matrilin-1 and Biglycan or Decorin Connect Collagen VI Microfibrils to Both Collagen II and Aggrecan. J Biol Chem 2003; 278: 37698-704. 59. Wagenera R et al.: The matrilins - adaptor proteins in the extracellular matrix. FEBS 2005; 579: 3323-9. 60. Shea MK et al.: Vitamin K and vitamin D status: association with inflammatory markers in the Framingham Offspring Study. Am J Epidemiol 2008; 167: 313-20. 61. El-Abbadi M, Giachelli CM: Mechanisms of vascular calcification. Advances in Chronic Kidney Disease 2007; 14: 54-66. 62. Schurgers LJ et al.: Oral anticoagulant treatment: friend and foe in cardiovascular disease? Blood 2004; 104: 3231-2. 63. Koos R et al.: Relation of oral anticoagulation to cardiac valvular and coronary calcium assessed by multislice spiral computed tomography. Am J Cardol 2005; 96: 747-9. 64. Hall JG, Pauli RM, Wilson KM: Maternal and fetal sequelae of anticoagulation during pregnancy. Am J Med 1980; 68: 122-40. 65. Howe AM et al.: Severe cervical dysplasia and nasal cartilage calcification following prenatal warfarin exposure. Am J Med Genet 1997; 71: 391-6. 66. Braam LA, Hoeks AP, Brouns F: Beneficial effects of vitamins D and K on the elastic properties of the vessel wall in postmenopausal women: a follow-up study. Thromb Haemost 2004; 91: 373-80. 67. Geleijnse JM, Vermeer C, Grobbee DE: Dietary intake of menaquinone is associated with a reduced risk of coronary heart disease: The Rotterdam Study. J Nutr 2004; 134: 3100-5. 68. Allison AC: The possible role of vitamin K deficiency in the pathogenesis of Alzheimer´s disease and in augmenting brain damage associated with cardiovascular disease. Med Hypotheses 2001; 57: 151-5. 69. Kohlmeier M, Salomon A, Saupe J, Shearer MJ: Transport of vitamin K to bone In humans. J Nutr 1996; 126 (suppl): 1192S-6S . 70. Cauley JA et al.: Apolipoprotein E Polymorphism: A New Genetic Marker of Hip Fracture Risk-The Study of Osteoporotic Fractures. J Bone Miner Res 1999; 14: 1175-81. 71. Thijssen HHW, Drittij-Reijnders MJ: Vitamin K distribution in rat tissue: dietary phylloquinone is a source of tissue menaquinone-4. Br J Nutr 1994; 72: 415-25. 72. Lev M, Milford AF: Effect of vitamin K depletion and restoration on sphingolipid metabolism in Bacteroides melaninogenicus. J Lipid Res 1972; 13: 364-70. 73. Sundaram KS, Lev Meir: Warfarin administration reduces synthesis of sulfatides and other sphingolipids in mouse brain. J Lipid Res 1998; 29: 1475-9. 74. Sundaram KS et al.: Vitamin K status influences brain sulfatide metabolism in young mice and rats. J Nutr 1996; 126: 2746-51. 75. Mitchell JS, Simon-Reuss I: Combination of some effects of x-radiation and a synthetic vitamin K substitute. Nature 1947; 160: 98-9. 76. Sasaki R et al.: DNA polymerase g inhibition by vitamin K3 induces mitochondria-mediated cytotoxicity in human cancer cells. Cancer Sci 2008; 99: 1040-8. 77. Nimptsch K, Rohrmann S, Linseisen J: Dietary intake of vitamin K and risk of prostate cancer in the Heidelberg cohort of the European Prospective Investigation into Cancer and Nutrition (EPIC-Heidelberg). Am J Clin Nutr 2008; 87: 985-92. 78. Noto V et al.: Effects of sodium ascorbate (vitamin C) and 2-methyl-1,4-naphthoquinone (vitamin K3) treatment on human tumor cell growth in vitro.Synergism of combined viatmin C and K3 action. Cancer 1989; 63: 901-6. 79. Sibayama-Imazu T et al.: Induction of apoptosis in PA-1 ovarian cancer cells by vitamin K2 is associated with an increased in the level of TR3/Nur77 and its accumulation in mitochondria and nuclei. J Cancer Res Clin Oncol 2008; 134: 803-12. 80. Wu Q et al.: Modulation of retinoic acid sensitivity in lung cancer cells through dynamic balance of orphan receptors nur77 and COUP-TF and their heterodimerization. EMBO J 1997; 16: 1656-69. 81. Habu D, Shiomi S, Tamori A: Role of vitamin K2 in the development of hepatocellular carcinoma in women with viral cirrhosis of the liver. JAMA 2004; 292: 358-61. 82. Kaneda M et al.: Vitamin K2 suppress malignancy of HuH7 hepatoma cells via inhibition of connexin 43. Cancaer Lett 2008; 263: 53-60. 83. Cummings SR, Melton LJ: Epidemiology and outcomes of osteoporotic fractures. Lancet 2002; 359: 1761-7. 84. Women Health Intitative study - USA 2006: N Engl J Med, Feb. 16. 85. Kaneki M et al.: Japanese fermented soybean food as the major determinant of large geographic difference in circulating levels of vitamin K2: possible implications for hip-fracture risk. Nutrition 2001; 17: 315-21.
