© Borgis - Postępy Nauk Medycznych 7/2012, s. 599-604
*Teresa Jackowska1, 2, Joanna Wójtowicz2
Rola hepcydyny w stanach zapalnych
The role of hepcidin in inflammation**
1Department of Paediatrics, Medical Centre of Postgraduate Education, Warsaw
Head of Department: prof. Teresa Jackowska, MD, PhD
2Clinical Department of Pediatrics, The Bielanski Hospital, Warsaw
Head of Department: prof. Teresa Jackowska, MD, PhD
Streszczenie
Niedawne odkrycie hepcydyny znacznie ożywiło badania nad gospodarką żelazem. Działanie hepcydyny, która hamuje wchłanianie i uwalnianie żelaza z komórek, wyjaśniło dotychczasową wiedzę na temat mechanizmów wchłaniania i spichrzania żelaza. Dzięki opisowi stymulacji stężenia hepcydyny stanem zapalnym i jej hamującego wpływu na stężenie żelaza w surowicy, udało się w pełni zrozumieć mechanizmy patologiczne związane między innymi z obniżeniem stężenia żelaza i niedokrwistości w czasie zakażeń i chorób przewlekłych.
W artykule przedstawiono podstawowe cechy hepcydyny, jej działanie, czynniki regulujące jej stężenie, a w szczególności rolę hepcydyny w stanach zapalnych. Przedstawiono potencjalne wykorzystanie wiedzy na temat hepcydyny w procesach diagnostyki różnicowej, np. w rozróżnieniu pomiędzy niedokrwistością chorób przewlekłych, a niedokrwistością z niedoboru żelaza. Ukazano perspektywiczne wykorzystanie substancji osłabiających działanie hepcydyny dla zmniejszenia stanów patologicznych związanych z zapaleniem.
Summary
The recent discovery of hepcidin has significantly intensified research on iron metabolism. Hepcidin, which inhibits iron intake and release from cells, has turned out to be a crucial player in the mechanisms of iron absorption and accumulation. Since hepcidin concentration is enhanced during inflammation, and hepcidin lowers the iron concentration itself, it is now more clear how pathologies like hypoferremia and anemia that accompany infections and chronic inflammation occur.
The key issues covered in the article are: basic facts about hepcidin, its physiological function and factors which determine its concentration, followed by an analysis of the role of hepcidin in inflammation. The paper also presents how knowledge about hepcidin may potentially facilitate the process of differential diagnosis, for example: iron-deficiency anemia versus anemia of chronic disease. A list of substances taken into account to be used as therapeutics that would reduce the influence of hepcidin on the iron concentration in chronic inflammation is presented at the end.

The history of discovering hepcidin
In 2000 the LEAP-1 (liver expressed antimicrobial peptide) molecule was isolated from human blood (1). A year later the same peptide was found in human urine, and was called hepcidin, due to the similarity of the amino acid sequence to murine hepcidin produced in the liver (2). Hepcidin was then qualified as belonging to the group of antibacterial peptides, alongside with defensins and protegrins, with regard to its structure that is rich in cysteines, as well as its antifungal and antibacterial activity in in vitro conditions. Thus, its name: hepcidin, meaning produced in the liver and of antibacterial activity (2).
However, it soon turned out that hepcidin is not merely another defensive protein that was discovered, but a central agent unifying the metabolic pathways regulating the iron concentration in the body. Its discovery significantly revived the research on iron metabolism in the last decade and gave a solid basis for using this knowledge in future clinical practice.
The activity of hepcidin
The basic activity of hepcidin is binding to ferroportin, the only protein described to date that transports iron from the cytoplasm to the extracellular space, e.g. to blood. As a result of the binding of hepcidin with ferroportin occurs an internalisation of ferroportin and its degradation, which in consequence leads to a reduction of iron concentration in serum (fig. 1). After the deactivation of ferroportin, iron is accumulated inside cells, e.g. the enterocytes, macrophages, hepatocytes and placenta cells (tab. 1) (3).
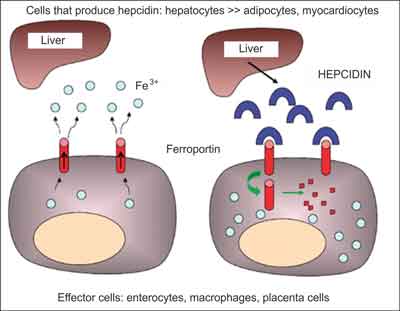
Fig. 1. Schema of the impact of hepcidin on a decreased iron absorption/release from effector cells (modified from (3)). The left schema presents the iron release by ferroportin. The right schema presents the activity of hepcidin, which binds to ferroportin, leading to its internalisaton and degradation, which in turn makes the transport of iron to blood impossible.
Table 1. Hepcidin activity.
| Hepcidin activity |
| Inhibiting the release of iron from enterocytes |
| Inhibiting the release of iron from hepatocytes |
| Inhibiting the release of iron from macrophages |
| Inhibiting the release of iron form placenta cells |
It has been empirically demonstrated that there is an inversely proportional relationship between the concentration of hepcidin and the concentration of iron in serum. Hepcidin was administered in three different doses to mice, and the observed decrease of iron concentration in serum was proportional to that dose (4). The only peptide active in this respect is the 25-amino acid sequence of hepcidin, as opposed to its metabolites, i.e. the 20- and 22-amino acid peptides. A similar activity is also not present in the precursor peptide for hepcidin i.e. prohepcidin, which was initially regarded as very promising, as it is easier to determine its concentration in body fluids (5).
It has also been demonstrated that there is an inverse relationship between hepcidin and iron concentration, in a clinical study with an iron overload, where the iron was administered orally to the participants. This caused an increased hepcidin concentration in serum (6). The results gave the basis to propose a study on hepcidin concentration as a reliable marker of iron metabolism, e.g. to predict the effectiveness of iron absorption when it is taken orally or to predict the response to treatment with ESA (erythropoesis stimulating agents) (7).
Understanding the metabolic pathways of hepcidin activity related to the effectiveness of iron absorption and accumulation has made it possible to explain the pathophysiological mechanisms of many diseases. Among them are hemochromatoses, where due to a mutation of the metabolic pathways an iron overload in tissue is caused (8) and in syderopenic anemia refractory to orally iron administration, where the iron absorption from the small intestine is inhibited through an excessive transcription of hepcidin (9).
Various mutations that condition the final increased or decreased activity of hepcidin have been described to date based on animal models and clinical cases. A severe iron overload of tissue was related e.g. to a mutation of the HAMP gene that encodes hepcidin in mice (10). Also, mutations of proteins that physiologically lead to an increase in hepcidin secretion (e.g. hemojuvelin knockout) cause a significant decrease in hepcidin concentration, and thus, an increased iron absorption and release (11). On the other hand, an overexpression of hepcidin causes animals to die shortly after birth due to severe anemia (12).
In humans, mutations of genes related to the regulation of hepcidin expression and activity in hemochromatoses have been described (8). In hepatocellular carcinoma, as well as in the Castelman disease, related to an excessive synthesis of interleukin 6 (IL-6), an overproduction of hepcidin was observed (13, 14). An increased or decreased activity of hepcidin may be connected with an abnormal structure of the ferroportin molecule, which affects the effectiveness of hepcidin binding with this protein, as well as the ultimate result of the regulation of iron metabolism.
Factors regulating hepcidin concentration
Four basic groups of factors regulate the hepcidin concentration in serum (tab. 2) (15). The factors responsible for a decreased hepcidin concentration are erythropoiesis and hypoxia. Both mechanisms condition an decreased hepcidin concentration, and thus, a decreased inhibition of iron absorption. Iron then becomes more accessible for the newly created erythrocytes.
Table 2. Four groups of factors that regulate hepcidin concentration (description and symbol explanation in the text).
| Factors regulating hepcidin concentration |
| | Impact on hepcidin
| Regulatory factors |
| Erythropoiesis | Decrease
↓ | GDF15, TWSG1, ESA
(unknown intracellularpathways) |
| Hypoxia | Decrease
↓ | Hypoxia, oxidative stress
(through the factors of HIF and CREBH) |
| Iron levelin serum | Increase
↑ | Transferrin-iron complex (Tf-Fe)
(by SMAD proteins) |
| Inflammation | Increase
↑ | interleukin-6
(through STAT proteins) |
Powyżej zamieściliśmy fragment artykułu, do którego możesz uzyskać pełny dostęp.
Mam kod dostępu
- Aby uzyskać płatny dostęp do pełnej treści powyższego artykułu albo wszystkich artykułów (w zależności od wybranej opcji), należy wprowadzić kod.
- Wprowadzając kod, akceptują Państwo treść Regulaminu oraz potwierdzają zapoznanie się z nim.
- Aby kupić kod proszę skorzystać z jednej z poniższych opcji.
Opcja #1
29 zł
Wybieram
- dostęp do tego artykułu
- dostęp na 7 dni
uzyskany kod musi być wprowadzony na stronie artykułu, do którego został wykupiony
Opcja #2
69 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 30 dni
- najpopularniejsza opcja
Opcja #3
129 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 90 dni
- oszczędzasz 78 zł
Piśmiennictwo
1. Krause A, Neitz S, Magert HJ et al.: LEAP-1, a novel highly disulfide-bonded human peptide, exhibits antimicrobial activity. FEBS Lett 2000; 480 (2-3): 147-150.
2. Park CH, Valore EV, Waring AJ, Ganz T: Hepcidin, a urinary antimicrobial peptide synthesized in the liver. J Biol Chem 2001; 276 (11): 7806-7810.
3. Bergamaschi G, Villani L: Serum hepcidin: a novel diagnostic tool in disorders of iron metabolism. Haematologica 2009; 94 (12): 1631-1633.
4. Rivera S, Nemeth E, Gabayan V et al.: Synthetic hepcidin causes rapid dose-dependent hypoferremia and is concentrated in ferroportin-containing organs. Blood 2005; 106 (6): 2196-2199.
5. Roe MA, Spinks C, Heath AL et al.: Serum prohepcidin concentration: no association with iron absorption in healthy men; and no relationship with iron status in men carrying HFE mutations, hereditary haemochromatosis patients undergoing phlebotomy treatment, or pregnant women. Br J Nutr 2007; 97 (3): 544-549.
6. Zimmermann MB, Troesch B, Biebinger R et al.: Plasma hepcidin is a modest predictor of dietary iron bioavailability in humans, whereas oral iron loading, measured by stable-isotope appearance curves, increases plasma hepcidin. Am J Clin Nutr 2009; 90 (5): 1280-1287.
7. Roe MA, Collings R, Dainty JR et al.: Plasma hepcidin concentrations significantly predict interindividual variation in iron absorption in healthy men. Am J Clin Nutr 2009; 89 (4): 1088-1091.
8. Malyszko J: Hemojuvelin: the hepcidin story continues. Kidney Blood Press Res 2009; 32 (2): 71-76.
9. Melis MA, Cau M, Congiu R et al.: A mutation in the TMPRSS6 gene, encoding a transmembrane serine protease that suppresses hepcidin production, in familial iron deficiency anemia refractory to oral iron. Haematologica 2008; 93 (10): 1473-1479.
10. Nicolas G, Bennoun M, Devaux I et al.: Lack of hepcidin gene expression and severe tissue iron overload in upstream stimulatory factor 2 (USF2) knockout mice. Proc Natl Acad Sci USA 2001; 98 (15): 8780-8785.
11. Ahmad KA, Ahmann JR, Migas MC et al.: Decreased liver hepcidin expression in the Hfe knockout mouse. Blood Cells Mol Dis 2002; 29 (3): 361-366.
12. Nicolas G, Bennoun M, Porteu A et al.: Severe iron deficiency anemia in transgenic mice expressing liver hepcidin. Proc Natl Acad Sci USA 2002; 99 (7): 4596-4601.
13. Chung A, Leo K, Wong G et al.: Giant hepatocellular adenoma presenting with chronic iron deficiency anemia. Am J Gastroenterol 2006; 101 (9): 2160-2162.
14. Arlet JB, Hermine O, Darnige L et al.: Iron-deficiency anemia in Castleman disease: implication of the interleukin 6/hepcidin pathway. Pediatrics 2010; 126 (6): e1608-e1612.
15. Nicolas G, Chauvet C, Viatte L et al.: The gene encoding the iron regulatory peptide hepcidin is regulated by anemia, hypoxia, and inflammation. J Clin Invest 2002; 110 (7): 1037-1044.
16. Tanno T, Bhanu NV, Oneal PA et al.: High levels of GDF15 in thalassemia suppress expression of the iron regulatory protein hepcidin. Nat Med 2007; 13 (9): 1096-1101.
17. Tanno T, Porayette P, Sripichai O et al.: Identification of TWSG1 as a second novel erythroid regulator of hepcidin expression in murine and human cells. Blood 2009; 114 (1): 181-186.
18. Ashby DR, Gale DP, Busbridge M et al.: Plasma hepcidin levels are elevated but responsive to erythropoietin therapy in renal disease. Kidney Int 2009; 75 (9): 976-981.
19. Vecchi C, Montosi G, Zhang K et al.: ER stress controls iron metabolism through induction of hepcidin. Science 2009; 325 (5942): 877-880.
20. Peyssonnaux C, Zinkernagel AS, Schuepbach RA et al.: Regulation of iron homeostasis by the hypoxia-inducible transcription factors (HIFs). J Clin Invest 2007; 117 (7): 1926-1932.
21. Schmidt PJ, Toran PT, Giannetti AM et al.: The transferrin receptor modulates Hfe-dependent regulation of hepcidin expression. Cell Metab 2008; 7 (3): 205-214.
22. Sheikh N, Dudas J, Ramadori G: Changes of gene expression of iron regulatory proteins during turpentine oil-induced acute-phase response in the rat. Lab Invest 2007; 87 (7): 713-725.
23. Kemna E, Pickkers P, Nemeth E et al.: Time-course analysis of hepcidin, serum iron, and plasma cytokine levels in humans injected with LPS. Blood 2005. 106 (5): 1864-1866.
24. Semrin G, Fishman DS, Bousvaros A et al.: Impaired intestinal iron absorption in Crohn’s disease correlates with disease activity and markers of inflammation. Inflamm Bowel Dis 2006; 12 (12): 1101-1106.
25. Malyszko J, Mysliwiec M: Hepcidin in anemia and inflammation in chronic kidney disease. Kidney Blood Press Res 2007; 30 (1): 15-30.
26. McClung JP, Karl JP: Iron deficiency and obesity: the contribution of inflammation and diminished iron absorption. Nutr Rev 2009; 67 (2): 100-104.
27. van Eijk LT, Kroot JJ, Tromp M et al.: Inflammation-induced hepcidin-25 is associated with the development of anemia in septic patients: an observational study. Crit Care 2011; 15 (1): R9.
28. de Mast Q, Syafruddin D, Keijmel S et al.: Increased serum hepcidin and alterations in blood iron parameters associated with asymptomatic P. falciparum and P. vivax malaria. Haematologica 2010; 95 (7): 1068-1074.
29. Cherian S, Forbes D, Cook A et al.: An Insight into the Relationship between Hepcidin, Anemia, Infections and Inflammatory Cytokines in Pediatric Refugees: A Cross-Sectional Study. PLoS ONE 2008; 3 (12): 1-8.
30. Ganz T: Iron in innate immunity: starve the invaders. Curr Opin Immunol 2009; 21 (1): 63-67.
31. Coyne D: Hepcidin: clinical utility as a diagnostic tool and therapeutic target. Kidney Int 2011; 80: 240-244.
32. Nemeth E, Valore EV, Territo M et al.: Hepcidin, a putative mediator of anemia of inflammation, is a type II acute-phase protein. Blood 2003; 101 (7): 2461-2463.
33. Theurl I, Aigner E, Theurl M et al.: Regulation of iron homeostasis in anemia of chronic disease and iron deficiency anemia: diagnostic and therapeutic implications. Blood 2009; 113 (21): 5277-5286.
34. Ganz T, Nemeth E: The Hepcidin-Ferroportin System as a Therapeutic Target in Anemias and Iron Overload Disorders. Hematology 2011, ASH, Updates on Disorders of Iron Utilisation and Distribution.
35. Pietrangelo A: Hepcidin in human iron disorders: therapeutic implications. J Hepatol 2010; 54 (1): 173-181.
36. Hashizume M, Uchiyama Y, Horai N et al.: Tocilizumab, a humanized anti-interleukin-6 receptor antibody, improved anemia in monkey arthritis by suppressing IL-6-induced hepcidin production. Rheumatol Int 2010; 30 (7): 917923.
37. Sasu BJ, Cooke KS, Arvedson TL et al.: Antihepcidin antibody treatment modulates iron metabolism and is effective in a mouse model of inflammation-induced anemia. Blood 2010; 115 (17): 3616-3624.
38. Kawabata H, Tomosugi N, Kanda J et al.: Anti-interleukin 6 receptor antibody tocilizumab reduces the level of serum hepcidin in patients with multicentric Castleman’s disease. Haematologica 2007; 92 (6): 857-858.
39. Zaritsky J, Young B, Gales B et al.: Reduction of serum hepcidin by hemodialysis in pediatric and adult patients. Clin J Am Soc Nephrol 2010; 5 (6): 1010-1014.
