© Borgis - Postępy Fitoterapii 3/2013, s. 182-190
*Katarzyna Sułkowska-Ziaja, Bożena Muszyńska, Halina Ekiert
Związki bezazotowe w owocnikach grzybów z gromady Basidiomycota
Non-nitrogen compounds in fruitbodies of Basidiomycota species
Katedra i Zakład Botaniki Farmaceutycznej, Uniwersytet Jagielloński, Collegium Medicum, Kraków
Kierownik Katedry i Zakładu: dr hab. Halina Ekiert, prof. nadzw. UJ
Summary
Basidiomycota fungi are an important source of biologically active compounds. Many of these components having different actions i.a. antiviral, bacterio- and fungistatic, antiparasitic as well as antitumor, anti-inflammatory, vasoprotective and hypoglycemic, were found in their fruiting bodies. In the presented elaboration we adopted frequently used division of fungal metabolites into two groups: nitrogenous and non-nitrogenous. Compounds occurring in Basidiomycota fungi and belonging to that second metabolite group are represented by carbohydrates, lipids, poliacetylens, poliketides, isoprenoids, sterols, organic acids and phenolic compounds. It is noteworthy that most of them are characterised by little or none toxicity. The best known fungal metabolites are polysaccharides, terpenoids (mono-, sesqui-, di-, tri and tetraterpens) and phenolic components. Amongst variety of well-known pharmacologically active fungal polysaccharides in clinical practice are used lentinan and krestin (i.e. PSK) and also shizophyllan.

Wstęp
Główne grupy biogenetyczne związków chemicznych grzybów, należących do gromady Basidiomycota można sklasyfikować następująco (1):
1. związki wywodzące się z metabolizmu podstawowego cukrów (monosacharydy, disacharydy, polisacharydy, alkohole cukrowe, czwartorzędowe zasady amoniowe),
2. związki powstałe w wyniku przemian aktywnego octanu (poliketydy, izoprenoidy, sterole),
3. związki powstałe z przemian kwasów tłuszczowych (poliacetyleny),
4. związki wywodzące się biogenetycznie z kwasu szikimowego (fenole, kwasy fenolowe),
5. związki powstałe w wyniku przemian aminokwasów (aminy, aminy toksyczne, alkaloidy, peptydy),
6. związki powstałe z przemian aminokwasów aromatycznych (alkaloidy sporyszu, związki indolowe).
W niniejszym opracowaniu przyjęto często spotykany podział metabolitów grzybowych na związki azotowe i bezazotowe. Związki bezazotowe występujące w gatunkach z gromady Basidiomycota reprezentowane są przez węglowodany, lipidy, poliacetyleny, poliketydy, izoprenoidy, sterole, kwasy organiczne, związki fenolowe (2).
Polisacharydy
W owocnikach grzybów występują nieliczne monosacharydy w postaci wolnej. Są to fruktoza, fukoza, galaktoza, glukoza, mannoza, ksyloza, sedoheptuloza (3). Jedynym znalezionym disacharydem w stanie wolnym jest trehaloza, zbudowana z dwóch cząsteczek glukozy połączonych wiązaniem O-glikozydowym, obficie występująca w rodzajach Boletus, Suillus i Cortinarius (4).
Grupą związków szeroko rozpowszechnioną wśród grzybów wyższych, o wyróżniającej się aktywności biologicznej, są polisacharydy. Aktywność biologiczna uwarunkowana jest ich strukturą chemiczną. Polisacharydy grzybowe reprezentowane są głównie przez glukany, ale mogą również mieć strukturę galaktanów, czy mannanów. Istotną rolę odgrywa typ wiązania glikozydowego (α lub β) oraz struktura przestrzenna cząsteczki polisacharydu. Aktywność biologiczną wykazują jedynie β glukany (5).
Masy cząsteczkowe osiągają wysokie wartości od 100 do 1000 kDa. Szczególnie aktywne są polisacharydy o budowie linearnej, bez długich łańcuchów bocznych, co wiąże się z ich łatwiejszą rozpuszczalnością, a tym samym łatwiejszą przyswajalnością. Najczęściej w głównym łańcuchu występują wiązania β(1→3), a rozgałęzienia tworzone są przez wiązania β(1→6). Znane są także polisacharydy mające dodatkowo fragment peptydowy.
Mechanizm przeciwnowotworowego działania polisacharydów polega na stymulacji określonych części składowych układu odpornościowego, głównie limfocytów T i B, makrofagów i komórek NK, do wydzielania interleukin. Ponieważ polisacharydy nie wykazywały aktywności cytotoksycznej lub cytostatycznej w testach in vitro w stosunku do komórek nowotworowych, przyjęto hipotezę, że związki te mogą działać pośrednio, aktywując układ immunologiczny, nie niszcząc bezpośrednio komórki zmienionej nowotworowo. Działając immunostymulująco, powodują zwiększenie naturalnej, nieswoistej odpowiedzi odpornościowej. Obecnie stosuje się je w połączeniu z innymi metodami zwalczania nowotworów, takimi jak radioterapia czy chemioterapia, po chirurgicznym usunięciu guza pierwotnego. Okazało się, że zwiększają one okres przeżywalności wśród chorych, hamując rozwój nowotworu i polepszając ogólny stan chorego (6).
Zainteresowanie substancjami cytotoksycznymi, zawartymi w grzybach przypada na początek lat 40. ubiegłego stulecia. Wyodrębniono wtedy polisacharydowy kompleks ze ściany komórkowej drożdży piekarskich Saccharomyces cerevisiae o właściwościach immunostymulujących. Po podaniu doustnym zaobserwowano aktywację makrofagów i stymulację układu siateczkowo-śródbłonkowego.
Oprócz właściwości przeciwnowotworowych związki te mogą wykazywać również inne działania biologiczne, np. przeciwwirusowe, przeciwbakteryjne, przeciwpasożytnicze, przeciwzapalne, hipotensyjne, hipoglikemiczne, czy naczynioochronne. Spośród szeregu znanych aktywnych farmakologicznie polisacharydów grzybowych, w praktyce klinicznej stosowane są głównie lentinan, krestin (tzw. PSK) i shizofyllan (7). Strukturę schizofyllanu podano na rycinie 1.
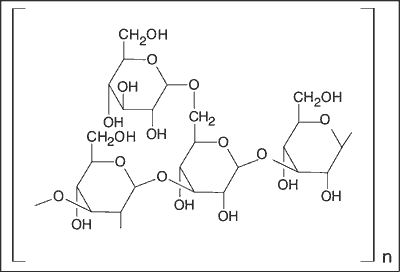
Ryc. 1. Budowa chemiczna schizofyllanu.
Lentinan został wyizolowany z owocników i kultur in vitro grzyba nadrzewnego Lentinula edodes. Chemicznie jest to β(1→3) glukan z rozgałęzieniami β(1→6) o masie cząsteczkowej 500 kDa, wykazujący strukturę prawoskrętnej helisy (8). Substancja ta uchodzi za najbardziej aktywną wśród poznanych polisacharydów o działaniu przeciwnowotworowym. Zapobiega powstawaniu zmian nowotworowych wywołanych przez karcynogeny chemiczne i poprzez wirusy, a także hamuje rozwój guzów allogenicznych i niektórych syngenicznych.
Polisacharyd ten stosowany jest najczęściej w leczeniu guzów litych żołądka, jelita grubego, piersi, płuc i w białaczce złośliwej. Prawdopodobny mechanizm jego działania polega na pobudzeniu limfocytów T, zwiększeniu wytwarzania interleukiny 1 i 3 oraz tlenku azotu przez komórki układu odpornościowego, stymulowaniu wydzielania CSF (Colony Stimulating Factor – czynnik stymulujący rozwój kolonii) i białek ostrej fazy oraz na bezpośrednim lub pośrednim (poprzez limfocyty T) wpływie na makrofagi. Stosowany jest w skojarzonym leczeniu łącznie z chemio- i radioterapią. Istotne jest to, że lentinan ma nieznaczne działanie uboczne, takie jak lokalne podrażnienia po wstrzyknięciu, czy sporadycznie gorączka i wymioty, lecz na ogół jest dobrze tolerowany przez organizm chorego (9).
Interesujące pod względem właściwości przeciwnowotworowych okazały się wyodrębnione z owocników Ganoderma lucidum frakcje polisacharydowe o charakterze glukuronoglukanów, ksyloglukanów, mannoglukanów i ksylomannoglukanów (10-12).
Również w Katedrze i Zakładzie Botaniki Farmaceutycznej UJ CM w Krakowie od wielu lat prowadzono badania nad polisacharydami izolowanymi z owocników grzybów wyższych.
Z gatunku Tylopilus felleus wyizolowano polisacharyd tylopilan o strukturze homoglukanu typu β(1→3) z rozgałęzieniami β(1→6), który w testach laboratoryjnych wykazywał działanie przeciwnowotworowe na komórki przeszczepialnego mięsaka Sarcoma-180 u myszy (13). Wykazał także właściwości immunomodulujące, wpływające na nieswoistą odporność komórkową, niektóre etapy swoistej odporności humoralnej oraz właściwości przeciwzapalne (14-16). Strukturę tylopilanu podano na rycinie 2.
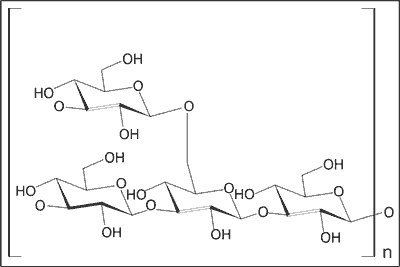
Ryc. 2. Budowa chemiczna tylopilanu.
Ponadto określono chemiczny charakter innych polisacharydów wyizolowanych z różnych gatunków z gromady Basidiomycota.
Do powszechnych składników grzybów należą alkohole cukrowe, przede wszystkim mannitol (17). W niektórych rodzajach, jak Russula czy Lactarius, zawartość mannitolu może dochodzić nawet do 20%. Inne alkoholocukry to sorbitol występujący w Suillus bovinus, ksylitol w Agaricus campestris oraz cykliczny m-inozytol obecny w Ramariopsis crocea (18).
W grzybach do tej pory nie znaleziono związków innych niż węglowodany zawierające połączenia glikozydowe, prawdopodobnie ze względu na brak systemów enzymatycznych umożliwiających proces glikozydacji.
Lipidy
Średnia całkowita zawartość lipidów w grzybach wynosi około 30%. Analiza jakościowa frakcji lipidowej wskazuje na obecność wolnych kwasów tłuszczowych, mono-, di- i triglicerydów, steroli oraz estrów steroli (19). W największej ilości występują wolne kwasy tłuszczowe oraz triglicerydy.
Zawartość wolnych kwasów tłuszczowych w grzybach waha się w granicach 20-30%. Zwraca uwagę znaczna zawartość kwasów tłuszczowych nienasyconych, które mogą stanowić nawet do 70% całkowitej zawartości kwasów tłuszczowych (20). Gatunkami o niewielkiej zawartości kwasów tłuszczowych są owocniki Agaricus bisporus i Pleurotus ostreatus, odpowiednio 0,3 i 0,4 g/100 g s.m. (21). Z kolei w gatunkach Cantharellus cibarius i Boletus edulis zawartość tych związków jest dużo wyższa i wynosi odpowiednio 2,6 oraz 3,6 g/100 g s.m. (22). Wśród związków nasyconych dominują kwasy: laurynowy, mirystynowy, palmitynowy, a wśród nienasyconych – oleinowy i linolowy. Na uwagę zasługuje ponadto obecność kwasu arachidonowego, znanego prekursora prostaglandyn (23).
Badania nad składem wolnych kwasów tłuszczowych w grzybach wskazują na biogenetyczne podobieństwo tych związków do świata roślin. W owocnikach Langermania gigantea 75% wszystkich kwasów tłuszczowych stanowi kwas linolowy. Inne dane literaturowe opisują kwas s-koriolinowy, którego strukturę podano na rycinie 3 oraz kwas linolenowy, związki wyizolowane z gatunku Pleurotus pulmonarius, wykazujące aktywność przeciwrobaczą (24).
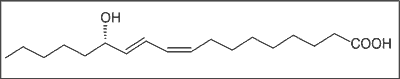
Ryc. 3. Budowa chemiczna kwasu s-koriolinowego wyizolowanego z Pleurotus pulmonarius.
Poliacetyleny
Powyżej zamieściliśmy fragment artykułu, do którego możesz uzyskać pełny dostęp.
Mam kod dostępu
- Aby uzyskać płatny dostęp do pełnej treści powyższego artykułu albo wszystkich artykułów (w zależności od wybranej opcji), należy wprowadzić kod.
- Wprowadzając kod, akceptują Państwo treść Regulaminu oraz potwierdzają zapoznanie się z nim.
- Aby kupić kod proszę skorzystać z jednej z poniższych opcji.
Opcja #1
29 zł
Wybieram
- dostęp do tego artykułu
- dostęp na 7 dni
uzyskany kod musi być wprowadzony na stronie artykułu, do którego został wykupiony
Opcja #2
69 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 30 dni
- najpopularniejsza opcja
Opcja #3
129 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 90 dni
- oszczędzasz 78 zł
Piśmiennictwo
1. Deacon J (red.). Fungal biology. Blackwell Pub, Malden 2006. 2. Kohlmünzer S, Grzybek J. Charakterystyczne składniki grzybów wielkoowocnikowych. Wiad Bot 1972; 16:35-176. 3. Khondkar P, Aidoo KE, Tester RF. Sugar profile of extracellular polysaccharides from different Tremella species. Int J Food Microbiol 2002; 79:121-9. 4. Wannet WJ, Aben EM, van der Drift C i wsp. Trehalose phosphorylase activity and carbohydrate levels during axenic fruiting in three Agaricus bisporus strains. Curr Microbiol 1999; 39:205-10. 5. Rop O, Mlcek J, Jurikova T. Beta-glucans in higher fungi and their health effects. Nutr Rev 2009; 67:624-31. 6. Komatsu N, Okubo S, Kikumoto S i wsp. Host mediated antitumor action of schizophyllan, a glucan produced by Schizophyllum commune. Gann 1969; 60:137-44. 7. Wasser SP. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl Microbiol Biotechnol 2002; 60:258-74. 8. Chihara G, Hamuro J, Maeda Y i wsp. Antitumor polysaccharides, lentinan and pachymaran. Saishin Igaku – Modern Med 1970; 25:1043-8. 9. Kurashige S, Akuzawa Y, Endo F. Effects of Lentinus edodes, Grifola frondosa and Pleurotus ostreatus administration on cancer outbreak, and activities of macrophages and lymphocytes in mice treated with a carcinogen, N-butyl-N-butanolnitrosoamine. Immunopharmacol Immunotoxicol 1997; 19:175-83. 10. Miyazaki T, Nishijima M. Studies on fungal polysaccharides. XXVII. Structural examination of a water-soluble, antitumor polysaccharide of Ganoderma lucidum, Chem Pharm Bull (Tokyo) 1981; 29:3611-6. 11. Kohda H, Tokumoto W, Sakamoto K i wsp. The biologically active constituents of Ganoderma lucidum (Fr.) Karst. Histamine release-inhibitory triterpenes. Chem Pharm Bull (Tokyo)1985; 33:1367-74. 12. Paterson RR. Ganoderma – a therapeutic fungal biofactory. Phytochem 2006; 67:1985-2001. 13. Kohlmünzer S, Grzybek J, Tanaka M. Antitumor and cytotoxic activity of polysaccharide from Tylopilus felleus. Planta Med 1980; 39:321-32. 14. Kohlmünzer S, Grzybek J, Wegiel J. Biological activity of polysaccharides from the mycelial culture of Tylopilus felleus (Bull. ex Fr.) P. Karst. Acta Pol Pharm 1992; 49:31-4. 15. Kohlmünzer S, Grzybek J, Molik-Wegiel J. Investigations on the biological activity of extracts of Tylopilus felleus (Bull. ex Fr.) P. Karst. by means of mycological tests. Pol J Pharmacol Pharm 1975; 27:95-9. 16. Kohlmünzer S, Quinn ML, Benoit PS i wsp. Antiinflammatory activity of Tylopilus felleus (Bull. ex Fr.) P. Karst. Pol J Pharmacol Pharm 1977; 29:539-41. 17. Ramstedt M, Jirjis R, Söderhäll K. Metabolism of mannitol in mycorrhizal and non-mycorrhizal fungi. New Phytol 1987; 105:281-7 18. Edmundowicz JM, Wriston JC. Mannitol dehydrogenase from Agaricus campestris. J Biol Chem 1963; 238:3539-41. 19. Feofilova EP. The lipids of mycelial fungi and the prospects for the development of microbial oleo-biotechnology. I. The lipids of mycelial fungi. Nauchnye Doki Vyss Shkoly Biol Nauk 1990; 11:5-25. 20. Brennan PJ, Griffin PF, Losel DM i wsp. The lipids of fungi. Prog Chem Fats Other Lipids 1974; 14:49-89. 21. Manzi P. Nutritional value of mushrooms widely consumed in Italy. Food Chem 2001; 73:321-5. 22. Karine P, Paul A, Andre G i wsp. Fatty acid composition of lipids from mushrooms belonging to the family Boletaceae. Mycol Res 2006; 110:1179-83. 23. Shier WT, Trotter JT. Stimulation of cell surface phospholipase A2 and prostaglandin synthesis in 3T2 mouse fibroblasts by phallolysin, a toxin from Amanita phalloides. Biochim Biophys Acta 1980; 619:235-46. 24. Zjawiony J. Biologically active compounds from Aphyllophorales (Polypore) Fungi. J Nat Prod 2004; 67:300-310. 25. Farrell IW, Keeping JW, Pellatt MG i wsp. Natural acetylenes. XLI. Polyacetylenes from fungal fruiting bodies. J Chem Soc 1973; 22:2642-3. 26. Anchel M. Structure of diatretyne 2, an antibiotic polyacetylenic nitrile from Clitocybe diatreta. Sci 1955; 121:607-8. 27. Anke T, Kupka J, Schramm G i wsp. Antibiotics from Basidiomycetes. X. Scorodonin, a new antibacterial and antifungal metabolite from Marasmius scorodonius (Fr.) Fr. J Antibiot (Tokyo) 1980; 33:463-7. 28. Ngai P, Zhao Z, Ng TB. Agrocybin, an antifungal peptide from the edible mushroom Agrocybe cylindracea. Peptides 2005; 26:191-6. 29. Sorg A, Siegel K, Bruckner R. Stereoselective syntheses of dihydroxerulin and xerulinic acid, anti-hypocholesterolemic dyes from the fungus Xerula melanotricha. Chemistry 2005; 11:1610-24. 30. Hu L, Tan JW, Liu JK. Chemical constituents of the basidiomycete Cortinarius umidicola. Z Naturforsch C. 2003; 58:659-62. 31. Weber W, Schu P, Anke T i wsp. Caloporoside, a new inhibitor of phospholipases C from Caloporus dichrous (Fr.) Ryv. J Antibiot (Tokyo) 1994; 47:1188-94. 32. Giannetti BM, Steglich W, Quack W i wsp. Antibiotics from Basidiomycetes, VI. Merulinic acids A, B, and C, new antibiotics from Merulius tremellosus and Phlebia radiata. Z Naturforsch C 1978; 33:807-16. 33. Liberra K, Jansen R, Wegner U i wsp. Fomecin B as a cytotoxic metabolite from the basidiomycete Tricholomopsis rutilans (Schaeff. ex. Fr.) Sing. Pharmazie 1995; 50:370-1. 34. Sontag B, Arnold N, Steglich W i wsp. Montadial A, a cytotoxic metabolite from Bondarzewia montana. J Nat Prod 1999; 62:1425-6. 35. Kohlmünzer S, Farmakognozja. Podręcznik dla studentów farmacji. Wyd Lek PZWL, Warszawa 1998. 36. Anchel M, Hervey A, Robbins WJ. Production of illudin M and of a fourth crystalline compound by Clitocybe illudens. Proc Natl Acad Sci 1952; 38:927-8. 37. Takeuchi T, Takahashi S, Iinuma H i wsp. Diketocoriolin B, an active derivative of coriolin B produced by Coriolus consors. J Antibiot (Tokyo) 1971; 24:631-5. 38. Takeuchi T, Iinuma H, Iwanaga J i wsp. Coriolin, a new Basidiomycetes antibiotic. J Antibiot (Tokyo) 1969; 22(5):215-7. 39. Luo DQ, Zhao LY, Shi YL i wsp. Velleratretraol, an unusual highly functionalized lactarane sesquiterpene from Lactarius vellereus. J Antibiot (Tokyo) 2009; 62:129-32. 40. Quack W, Anke T, Oberwinkler F i wsp. Antibiotics from Basidiomycetes. V. Merulidial, a new antibiotic from the Basidiomycete Merulius tremellosus Fr. J Antibiot (Tokyo) 1978; 31:737-41. 41. Kozova J, Rehacek Z. Antibiotics of Flammulina velutipes cultivated in submerged culture. Folia Microbiol (Praha). 1967; 12:567-8. 42. Stadler M, Anke H. New nematicidal and antimicrobial compounds from the basidiomycete Cheimonophyllum candidissimum (Berk & Curt.) sing. I. Producing organism, fermentation, isolation, and biological activities. J Antibiot (Tokyo) 1994; 47:1284-9. 43. Kavanagh F. Antibiotics substances from Basidiomycetes. Proc. NAS 1951; 570-4. 44. Allbutt AD, Ayer WA, Brodie HJ i wsp. Cyathin, a new antibiotic complex produced by Cyathus helenae. Can J Microbiol 1971; 17:1401-7. 45. Kenmoku H, Shimai T, Toyomasu T i wsp. Erinacine Q, a new erinacine from Hericium erinaceum, and its biosynthetic route to erinacine C in the basidiomycete. Biosci Biotechnol Biochem 2002; 66:571-5. 46. Paterson RR. Ganoderma – a therapeutic fungal biofactory. Phytochem 2006; 67:1985-2001. 47. Kamo T, Asanoma M, Shibata H i wsp. Anti-inflammatory lanostane-type triterpene acids form. J Nat Prod 2003; 66:1104-6. 48. Kahlos K, Hiltunen R. Triterpenes and sterols in Inonotus radiatus. Planta Med 1988; 54:579. 49. Kubo I, Matsumoto A, Kozuka M i wsp. Calmodulin inhibitors from the bitter mushroom Naematoloma fasciculare (Fr.) Karst. (Strophariaceae) and absolute configuration of fasciculols. Chem Pharm Bull (Tokyo) 1985; 33:3821-5. 50. Anke T, Werle A, Zapf S i wsp. Favolon, a new antifungal triterpenoid from a Favolaschia species. J Antibiot 1995; 48:725-6. 51. Muszyńska B, Sułkowska-Ziaja K, Ekiert H. An antioxidant in fruiting bodies and in mycelia from in vitro cultures of Calocera viscosa (Basidiomycota) – preliminary results. Acta Pol Pharm 2012; 69:135-8. 52. Marino A, Mustacchi S, Puntillo D i wsp. Sterols from Basidiomycetes. Boll Soc Ital Biol Sper 1985; 61:171-4. 53. Mattila P. Sterol and vitamin D2 contents in some wild and cultivated mushrooms. Food Chem 2002; 76:293-8. 54. Kahlos K, Kangas L, Hiltunen R. Ergosterol peroxide, an active compound from Inonotus radiatus. Planta Med 1989; 55:389-90. 55. Yuan JP, Kuang HC, Wang JH i wsp. Evaluation of ergosterol and its esters in the pileus, gill, and stipe tissues of agaric fungi and their relative changes in the comminuted fungal tissues. Appl Microbiol Biotechnol 2008; 80:459-65. 56. Weete JD, Kelley WD, Hollis CA. Mycelial lipids as an aid in identifying rust fungi in culture – Cronartium fusiforme. Can J Microbiol 1979; 25:1481-3. 57. Takao S. Organic acid production by Basidiomycetes. I. Screening of acid-producing strains. Appl Microbiol 1965; 13:732-37. 58. Ribeiro B, Rangel J, Valentao P i wsp. Contents of carboxylic acids and two phenolics and antioxidant activity of dried Portuguese wild edible mushrooms. J Agric Food Chem 2006; 54:8530-7. 59. Carminati GM, Spina G. Pharmacological properties of agaric acid. Boll Chim Farm 1968; 107:249-51. 60. Lomascolo A. Basidiomycetes as new biotechnological tools to generate natural aromatic flavours for the food industry. Trends Biotechnol 1996; 17:282-9. 61. Zhou X, Gong Z, Su Y i wsp. Cordyceps fungi: natural products, pharmacological functions and developmental products. J Pharm Pharmacol 2009; 61:279-91. 62. Mattila P, Konko K, Eurola M i wsp. Contents of vitamins, mineral elements, and some phenolic compounds in cultivated mushrooms. J Agric Food Chem 2001; 49:2343-8. 63. Barros L, Duenas M, Ferreira IC i wsp. Phenolic acids determination by HPLC-DAD-ESI/MS in sixteen different Portuguese wild mushrooms species. Food Chem Toxicol 2009; 47:1076-9. 64. Barros L, Baptista P, Correia DM i wsp. Effects of conservation treatment and cooking on the chemical composition and antioxidant activity of Portuguese wild edible mushrooms. J Agric Food Chem 2007; 55:4781-8. 65. Lu X, Chen U, Dong P i wsp. Phytochemical characteristics and hypoglycaemic activity of fraction from mushroom Inonotus obliquus. J Sci Food Agric 2009; 90:276-80.
