© Borgis - Medycyna Rodzinna 4/2016, s. 223-227
Katarzyna Osmańska1, Ewelina Łazarczyk1, Barbara Mucha1, Sylwia Stąpor2
Znaczenie genu ETV6 w hematopoezie i leukemogenezie
The significance of ETV6 gene in hematopoiesis and leukemogenesis
1Katedra i Zakład Genetyki Klinicznej, Wydział Lekarski, Uniwersytet Mikołaja Kopernika – Collegium Medicum w Bydgoszczy
2Katedra i Klinika Hematologii, Nowotworów Krwi i Transplantacji Szpiku, Uniwersytet Medyczny im. Piastów Śląskich we Wrocławiu
Summary
Chromosomal aberrations and gene mutations play important role in hematological malignancies. Translocations involving 12p13 region are one of the most commonly observed chromosomal abnormalities in leukemias, myelodysplastic syndromes and myeloproliferative neoplasms. ETV6 (ETS translocation variant gene 6) gene, located in 12p13 band, is a member of ETS (E26 transformation-specific) transcription factors family. Members of ETS family are involved in angiogenesis and hematopoiesis, as well as growth and differentiation of cells. ETV6 is a strong transcriptional repressor, in which three domains can be distinguished: HLH (helix-loop-helix), ETS and internal domain. 48 chromosomal bands involved in ETV6 translocations, insertions and inversions have been found so far. Moreover, 30 ETV6 partner genes have been revealed. They belong to different classes: receptor and non-receptor tyrosine kinase genes, transcription factor genes, homeobox genes and others. ETV6-RUNX1 (TEL-AML1) is the most common ETV6 rearrangement. It results from t(12;21)(p13;q22).

Wstęp
Do głównych zadań układu krwiotwórczego należy stałe wytwarzanie elementów morfotycznych krwi. Aby komórki krwi wytwarzane w procesie hematopoezy mogły wypełniać swoją rolę, potrzebnych jest wiele czynników, m.in. prawidłowa funkcja genomu.
Białaczki są chorobami rozrostowymi, nowotworowymi, których przyczyna nie jest jednoznacznie określona. Ich cechą charakterystyczną jest klonalny rozrost wielopotencjalnej komórki macierzystej. Obecnie stosowaną klasyfikacją nowotworów układu krwiotwórczego jest klasyfikacja WHO z 2008 roku.
Ważną rolę w nowotworach hematologicznych odgrywają aberracje chromosomowe i mutacje genowe. Pierwszą zidentyfikowaną aberracją chromosomową była zrównoważona wzajemna translokacja pomiędzy chromosomami par 9 i 22 [t(9;22)(q34;q11.2)], skutkująca powstaniem dwóch genów fuzyjnych BCR-ABL na der(22) oraz ABL-BCR na der(9), opisana w roku 1960 przez Petera Nowella i Davida Hungerforda. W kolejnych latach odkryto wiele innych translokacji, wyznaczających fenotyp komórek białaczkowych i przebieg kliniczny określonego typu białaczki (1). Translokacje angażujące region 12p13 są jednymi z najczęściej obserwowanych zmian chromosomowych w białaczkach i zespołach mielodysplastycznych. W regionie tym znajduje się gen ETV6 (ang. ETS-translocation variant gene 6), należący do rodziny czynników transkrypcyjnych ETS (ang. E26 transformation-specific), do której zalicza się aktywatory i represory transkrypcji. ETV6 bierze udział w angiogenezie, prawidłowym przebiegu hematopoezy, a także w procesie wzrostu i różnicowania się komórek. Jest także zaangażowany w różnorodne rearanżacje prowadzące do rozwoju nowotworów układu krwiotwórczego. Wysoka częstość, z jaką gen ETV6 angażowany jest w procesy leukemogenezy, wynika z dużej liczby genów partnerskich, przy współudziale których doprowadza on do transformacji komórek prawidłowych w komórki białaczkowe (2-4).
Charakterystyka genu ETV6
Gen ETV6 zlokalizowany jest na chromosomie 12, w regionie p13.1 (1, 3, 5). Odkrywcami genu byli Golub i Gilliland, którzy w 1994 roku ogłosili wyniki swoich badań nad translokacją t(5;12)(q33;p13). W wyniku tej translokacji, która opisana została po raz pierwszy u chorego cierpiącego na przewlekłą białaczkę mielomonocytową (ang. chronic myelomonocytic leukemia – CMML), dochodzi do powstania genu fuzyjnego ETV6-PDGFRβ (3).
Pierwotnie gen ETV6 nosił nazwę TEL (ang. translocation ETS leukemia). Później, aby uniknąć niejasności (skrót „tel” używa się do określenia telomerów chromosomów), zmieniono jego nazwę na ETV6. Obecnie w literaturze częściej stosowana jest nowsza nazwa (1, 3, 6). ETV6 należy do rodziny czynników transkrypcyjnych – ETS. Białka ETS zostały po raz pierwszy opisane w ptasim wirusie erytroblastozy – E26. Rodzina tych białek uważana jest za jedną z największych rodzin czynników transkrypcyjnych, o zróżnicowanych funkcjach. Można w niej wyróżnić zarówno aktywatory, jak i represory transkrypcji. Genami docelowymi rodziny ETS są geny odpowiedzialne za procesy różnicowania, proliferacji, hematopoezy, apoptozy, angiogenezy i nabywania przez komórki właściwości inwazyjnych (1, 4, 7).
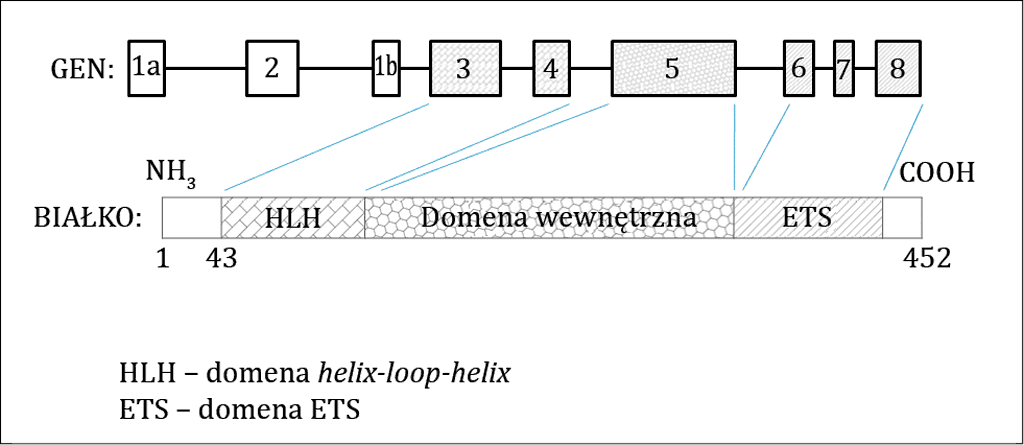
ETV6 zbudowany jest z ośmiu eksonów, których łączna długość wynosi ok. 40 kpz (kpz – kilo par zasad = tysiąc par zasad) (8). Eksony 3 i 4 kodują domenę HLH (ang. helix-loop- -helix – helisa-pętla-helisa), zwaną inaczej domeną B. Domena ETS (domena wiążąca DNA) kodowana jest przez eksony 6, 7 i 8. Źródła literaturowe wyróżniają dodatkowo domenę wewnętrzną (ang. internal domain), kodowaną przez ekson 5. Pierwsze dwa introny genu, mierzące 100 i 82 kpz, są najdłuższymi intronami w całym genie. Długość pozostałych pięciu intronów (od intronu 3 do 7) waha się w granicach 1,3-15 kpz. Sekwencja zwana wyspą CpG zlokalizowana jest na końcu 5’ genu (ryc. 1). Obecność sekwencji tego typu w formie niezmienionej (niezmetylowanej) jest istotna dla ekspresji genu (1, 9). Najbardziej zbliżony do genu ETV6 pod względem budowy jest gen ETS1 (ang. v-ets erythroblastosis virus E26 oncogene homolog 1 (avian)), należący do tej samej rodziny czynników transkrypcyjnych. Obydwa geny zbudowane są z ośmiu eksonów i mają długi pierwszy intron (10).

Ryc. 1. Struktura genu oraz białka ETV6. GEN – prostokąty oznaczają kolejne eksony (od 1 do 8). Odcinki proste oznaczają introny. Poszczególne eksony kodujące odpowiednie domeny (eksony 3 i 4 – domena HLH; ekson 5 – domena wewnętrzna, eksony 6, 7 i 8 – domena ETS) oznaczono różnymi teksturami. BIAŁKO – poszczególne domeny (domena HLH, domena wewnętrzna oraz domena ETS) oznaczono różnymi teksturami (analogicznie do oznaczeń zastosowanych do eksonów je kodujących). Liczby 1, 43 i 452 oznaczają kolejne aminokwasy w strukturze białka
Struktura białka ETV6
Na białko ETV6 składają się 452 aminokwasy. ETV6 jest jądrową fosfoproteiną, której ekspresja zachodzi w zdrowych tkankach pochodzących z układu krwiotwórczego i spoza tego układu, jak również w wielu nowotworach. W obrębie ETV6 znajdują się trzy miejsca fosforylacji kinaz aktywowanych mitogenami (ang. mitogen activated protein kinase – MAPK; kinazy aktywowane mitogenami). Końce 5’ i 3’ białka ETV6 zbudowane są analogicznie do końców pozostałych białek z rodziny ETS. Fosfoproteina ETV6 może występować w komórkach w jednej z dwóch izoform. Masy cząsteczkowe izoform wynoszą odpowiednio 50 i 57 kDa. Izoformy ETV6 powstają na drodze alternatywnego splicingu pre-mRNA – mechanizmu zwiększającego różnorodność białek. Izoforma o masie 57 kDa zawiera o 42 aminokwasy więcej od izoformy krótszej. Jest to wynik obecności dwóch kodonów startowych: Met1 i Met43. Pierwszy kodon startowy zlokalizowany jest na początku genu w obrębie eksonu 1a. Natomiast drugi, alternatywny kodon startowy znajduje się na początku eksonu 1b zlokalizowanego tuż przed eksonem 3. Rozpoczęcie translacji od drugiego kodonu startowego prowadzi do utraty jednego z miejsc fosforylacji MAPK, zlokalizowanego w obrębie dwudziestego drugiego aminokwasu kodującego serynę (Ser22). Różna liczba miejsc fosforylacji MAPK w poszczególnych izoformach jest odpowiedzialna za różnice w ich aktywności (2, 11).
Powyżej zamieściliśmy fragment artykułu, do którego możesz uzyskać pełny dostęp.
Mam kod dostępu
- Aby uzyskać płatny dostęp do pełnej treści powyższego artykułu albo wszystkich artykułów (w zależności od wybranej opcji), należy wprowadzić kod.
- Wprowadzając kod, akceptują Państwo treść Regulaminu oraz potwierdzają zapoznanie się z nim.
- Aby kupić kod proszę skorzystać z jednej z poniższych opcji.
Opcja #1
29 zł
Wybieram
- dostęp do tego artykułu
- dostęp na 7 dni
uzyskany kod musi być wprowadzony na stronie artykułu, do którego został wykupiony
Opcja #2
69 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 30 dni
- najpopularniejsza opcja
Opcja #3
129 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 90 dni
- oszczędzasz 78 zł
Piśmiennictwo
1. de Braekeleer E, Douet-Guibert N, Morel F et al.: ETV6 fusion genes in hematological maligancies: A review. Leuk Res 2012; 36: 945-961. 2. Bohlander SK: ETV6: A versatile player in leukemogenesis. Semin Cancer Biol 2005; 15: 162-174. 3. Haferlach C, Bacher U, Schnittger S et al.: ETV6 rearrangements are recurrent in myeloid malignancies and are frequently associated with other genetic events. Genes Chromosomes Cancer 2012; 51: 328-337. 4. Kralik JM, Kranewitter W, Boesmueller H et al.: Characterization of a newly identified ETV6-NTRK3 fusion transcript in acute myeloid leukemia. Diagn Pathol 2011; 6: 19-23. 5. Zaliova M, Meyer C, Cario G et al.: TEL/AML1-positive patients lacking TEL exon 5 resemble canonical TEL/AML1 cases. Pediatr Blood Cancer 2011; 56: 217-225. 6. Barjesteh van Waalwijk van Doorn-Khosrovani S, Spensberger D, de Knegt Y et al.: Somatic heterozygous mutations in ETV6 (TEL) and frequent absence of ETV6 protein in acute myeloid leukemia. Oncogene 2005; 24: 4129-4137. 7. Lesiak K, Sztiller-Sikorska M, Czyż M: Czynniki transkrypcyjne w powstawaniu i progresji czerniaka. Postepy Hig Med Dosw 2007; 61: 576-595. 8. Sawińska M, Ładoń D: Mechanism, detection and clinical significance of the reciprocal translocation t(12;21)(p12;q22) in the children suffering from acute lymphoblastic leukaemia. Leuk Res 2004; 28: 35-42. 9. Eguchi-Ishimae M, Eguchi M, Maki K et al.: Leukemia-related transcription factor TEL/ETV6 expands erythroid precursors and stimulates hemoglobin synthesis. Cancer Sci 2009; 100: 689-697. 10. Beans M, Peeters P, Guo C et al.: Genomic organization of TEL: the human ETS-variant gene 6. Genome Res 1996; 6: 404-413. 11. van Rompaey L, Potter M, Adams C, Grosveld G: Tel induces a G1 arrest and suppresses Ras-inducted transformation. Oncogene 2000; 19: 5244-5250. 12. Wang LC, Swat W, Fujiwara Y et al.: The TEL/ETV6 gene is required specifically for hematopoiesis in bone marrow. Genes Dev 1998; 12: 2392-2402. 13. Hock H, Meade E, Medeiros S et al.: Tel/Etv6 is an essential and selective regulator of adult hematopoietic stem cell survival. Genes Dev 2004; 18: 2336-2341. 14. Montpetit A, Wilson MD, Chevrette M et al.: Analysis of the conservation of synteny between Fugu and human chromosome 12. BMC Genomics 2003; 4: 30-37. 15. Deves C, Renck D, Garicochea B et al.: Analysis of select members of the E26 (ETS) transcription factors family in colorectal cancer. Virchows Arch 2011; 458: 421-430. 16. Nand R, Bryke C, Kroft SH et al.: Myeloproliferative disorder with eosinophilia and ETV6-ABL gene rearrangement: Efficacy of second-generation tyrosine kinase inhibitors. Leuk Res 2009; 33: 1144-1146. 17. Gancheva K, Virchis A, Howard-Reeves J et al.: Myeloproliferative neoplasm with ETV6-ABL1 fusion: a case report and literature review. Mol Cytogenet 2013; 6: 39-48. 18. Gandemer V, Rio AG, de Tayrac M et al.: Five distinct biological processes and 14 differentially expressed genes characterize TEL/AML1-positive leukemia. BMC Genomics 2007; 8: 385-399. 19. Lee DS, Kim YR, Cho HK et al.: The presence of TEL/AML1 rearrangement and cryptic deletion of the TEL gene in adult acute lymphoblastic leukemia (ALL). Cancer Genet Cytogenet 2005; 162: 176-178. 20. Ellinghaus E, Stanulla M, Richter G et al.: Identification of germline susceptibility loci in ETV6-RUNX1-rearranged childhood acute lymphoblastic leukemia. Leukemia 2012; 26: 902-909. 21. von Goessel H, Jacobs U, Semper S et al.: Cluster analysis of genomic ETV6-RUNX1 (TEL-AML1) fusion sites in childhood acute lymphoblastic leukemia. Leuk Res 2009; 33: 1082-1088. 22. Al-Shehhi H, Konn ZJ, Schwab CJ et al.: Abnormalities of the der(12)t(12;21) in ETV6-RUNX1 acute lymphoblastic leukemia. Genes Chromosomes Cancer 2013; 52: 202-213. 23. Fischer M, Schwieger M, Horn S et al.: Defining the oncogenic function of the TEL/AML1 (ETV6/RUNX1) fusion protein in a mouse model. Oncogene 2005; 24: 7579-7591. 24. Zelent A, Greaves M, Enver T: Role of the TEL-AML1 fusion gene in the molecular pathogenesis of childhood acute lymphoblastic leukaemia. Oncogene 2004; 23: 4275-4283. 25. Kantner HP, Warsch W, Delogu A et al.: ETV6/RUNX1 induces reactive oxygen species and drives the accumulation of DNA damage in B cells. Neoplasia 2013; 15: 1292-1300. 26. Balatzenko G, Guenova M, Kalinova I et al.: Simultaneous occurrence of ETV6-RUNX1 and BCR-ABL1 (e1a2) transcripts in a child with B-cell acute lymphoblastic leukemia. Cancer Genet 2013; 206: 97-101. 27. Zakaria Z, Ahid MFM, Ismail A et al.: Chromosomal aberrations in ETV6/RUNX1-positive childhood acute lymphoblastic leukemia using 244K oligonucleotide array comparative genomic hybridization. Mol Cytogenet 2012; 5: 41-46. 28. Stams WAG, Beverloo HB, den Boer ML et al.: Incidence of additional genetic changes in the TEL and AML1 genes in DCOG and COALL-treated t(12;21)-positive pediatric ALL, and their relation with drug sensitivity and clinical outcome. Leukemia 2006; 20: 410-416. 29. Raynaud S, Cave H, Baens M et al.: The 12;21 translocation involving TEL and deletion of the other TEL allele: two frequently associated alterations found in childhood acute lymphoblastic leukemia. Blood 1996; 87: 2891-2899. 30. Sevilla DW, Nandulax SV, Colovai AI et al.: Diffuse large B-cell lymphoma with TEL/ETV6 translocation. Hum Pathol 2009; 40: 588-593. 31. Uphoff CC, MacLeod RAF, Denkmann SA et al.: Occurrence of TEL-AML1 fusion resulting from (12;21) translocation in human early B-lineage leukemia cell lines. Leukemia 1997; 11: 441-447.