© Borgis - Postępy Nauk Medycznych 4b/2015, s. 16-22
*Sylwia Jarzynka1, 2, Ewa Swoboda-Kopeć2, Maria Dąbkowska3, Ewa Augustynowicz-Kopeć1
Fenotypowa i genotypowa charakterystyka szczepów Candida spp. izolowanych od pacjentów z podejrzeniem fungemii odcewnikowych
Phenotypic and genotypic characteristic of Candida spp. strains isolated from patients with suspected catheter-related bloodstrem fungal infections
1Zakład Mikrobiologii, Instytut Gruźlicy i Chorób Płuc, Warszawa
Kierownik Zakładu: prof. dr hab. n. med. Ewa Augustynowicz-Kopeć
2Zakład Mikrobiologii, Centralny Szpital Kliniczny, Warszawa
P.o. Kierownika Zakładu: dr hab. n. med. Marta Wróblewska
3Katedra i Zakład Mikrobiologii, Warszawski Uniwersytet Medyczny
Kierownik Zakładu: prof. nadzw. dr hab. med. Grażyna Młynarczyk
Streszczenie
Wstęp. W ciągu ostatnich kilkudziesięciu lat odnotowuje się znaczny wzrost zakażeń grzybiczych u ludzi. Układowe zakażenia szczepami Candida spp. mogą przebiegać z rozsiewem hematogennym grzybów (fungemią).
Cel pracy. Celem pracy była charakterystyka fenotypowa i genotypowa 90 szczepów Candida spp. izolowanych z materiałów klinicznych pobieranych od chorych z podejrzeniem fungemii odcewnikowych.
Materiał i metody. Charakterystyka fenotypowa Candida spp. obejmowała ocenę aktywności enzymatycznej szczepów oraz ocenę ich zdolności do tworzenia struktury biofilmu. Charakterystyka genotypowa szczepów Candida spp. opierała się na wykrywaniu genów kodujących proteazy Sap i Yps.
Wyniki. Z materiałów klinicznych wyhodowano szczepy Candida należące do gatunków: C. albicans – 32,2%, C. parapsilosis – 30%, C. glabrata – 24,5%, C. krusei – 7,8%, C. tropicalis – 3,3% oraz C. guilliermondii – 2,2%. Szczepy C. albicans wykazywały odwrotną zależność między aktywnością proteaz a wzrostem w postaci biofilmu. Szczepy C. parapsilosis wykazywały wyższą zdolność tworzenia biofilmu w porównaniu do izolatów C. albicans.
Wnioski. Efg1 oraz N-acetylo-β-glukozaminidaza (NAG) mogą stanowić istotne czynniki zjadliwości szczepów C. albicans w aspekcie zdolności tworzenia biofilmu. W badaniach obserwowano zmniejszoną rolę proteaz aspartylowych (Sap) w wirulencji szczepów C. albicans.
Summary
Introduction. In the XXc. the high level of fungal infections is observed. Systemic fungal infections were associated with bloodstream candidiosis. Catheter-related bloodstream fungal infection is a haematogenous spread of fungi associated with vascular catheter colonization.
Aim. Aim of this work was to analyze the phenotypic and genotypic features of fungi isolated from clinical samples from patients with suspected catheter-related bloodstream fungal infection.
Material and methods. These studies are based on phenotypic methods, such as enzymatic activity and the ability to biofilm production. The genotypic evaluation were used to search for genes encoding virulence factors of fungi like Sap and Yps.
Results. The major etiological factors of catheter-related bloodstream fungal infections were species: C. albicans – 32.2%, C. parapsilosis – 30% and C. glabrata – 24.5%. An inverse relationship between the activity of proteases and growth of strains in the biofilm structure in the C. albicans strains were shown. Maximum capacity to grow the biofilm was characteristic for C. parapsilosis strains – 48%.
Conclusions. The presence of the EFG1 gene and NAG enzyme suggests ability to produce invasive hyphal forms, especially in aspect of biofilm formation. We observed lower role of Sap in C. albicans virulence.

Wstęp
W ciągu ostatnich kilkudziesięciu lat odnotowuje się znaczny wzrost zakażeń grzybiczych u ludzi. Dotyczy to zwłaszcza środowiska szpitalnego w grupie osób z obniżoną odpornością. Większość zakażeń grzybiczych stanowią kandydozy. Układowe zakażenia szczepami Candida spp. mogą przebiegać z rozsiewem hematogennym grzybów (fungemią). Od 40 do 60% wszystkich fungemii stanowią zakażenia odcewnikowe związane z kolonizacją cewników naczyniowych (obwodowych i centralnych) przez szczepy Candida spp. C. albicans to gatunek dominujący (44,8%) wśród grzybiczych czynników etiologicznych fungemii odcewnikowych (1). Kolejne gatunki grzybów izolowane z zakażeń to C. glabrata (28,1%) oraz C. parapsilosis (10,4%) (1). Diagnostyka laboratoryjna gatunków innych niż Candida albicans jest długotrwała, ze względu na ich zróżnicowanie morfologiczne, specyficzne cechy biochemiczne oraz zmniejszoną wrażliwość na lek pierwszego rzutu – fulkonazol, a także echinokandyny (2).
Na chorobotwórczość grzybów mają wpływ: cechy fenotypowe i genotypowe, stopień zjadliwości, wrażliwość na chemioterapeutyki oraz przebieg zakażenia. Do czynników wirulencji grzybów należy zaliczyć: dimorfizm, phenotypic switching, zdolności adhezyjne, tworzenie biofilmu, signaling i quorum sensing oraz aktywność enzymatyczną. Grzyby wykazują zdolność wzrostu w różnych formach morfologicznych. Szczepy Candida spp. rosną w formie pojedynczych komórek – blastospor i/lub tworząc pseudostrzępki posiadające zdolność penetrowania w głąb tkanek gospodarza (3). Przemiana morfologiczna grzybów jest poprzedzona wieloma zmianami biochemicznymi, fizjologicznymi i strukturalnymi zachodzącymi w komórce grzyba. Związana jest z reorganizacją cytoszkieletu, poprzez zwiększenie zawartości chityny w strzępkach oraz z ekspresją specyficznych białek strukturalnych, enzymatycznych i regulatorowych. Do białek regulatorowych należą m.in.: Cph1, Cst20, Hst7, Cek1, Efg1, Rim101, Prr2, Ras, Tpk2, Tup1p, Rbf1, Nrg1, Rfg1, Flo8, Czf1 i Tec1 (3, 4). Zdolność dimorfizmu jest regulowana nie tylko przez temperaturę, ale także przez wiele innych czynników, takich jak stres osmotyczny, dostępność substancji odżywczych i CO2 czy pH środowiska (ryc. 1).

Ryc. 1. Czynniki regulujące przemianę morfologiczną C. albicans [wg (4), zmodyfikowano].
Na stopień zjadliwości grzybów drożdżopodobnych ma wpływ także różnorodność fenotypowa (Phenotypic switching), czyli zdolność występowania różnych form morfologicznych w obrębie tego samego gatunku. Zmiana fenotypu kolonii wiąże się z odmiennymi właściwościami biochemicznymi, metabolicznymi, chorobotwórczymi. Szczepy Candida spp. przechodzące przemianę morfologiczną są znacznie częściej przyczyną kandydoz przebiegających z zakażeniem krwi i/lub infekcją odcewnikową (4). Kolejnym czynnikiem wirulencji szczepów Candida spp. jest zdolność adhezji. Adhezyny pozwalają na adaptację komórek grzybów do różnych środowisk, m.in. do powierzchni komórek nabłonkowych, błony komórkowej, wewnątrzkomórkowych białek macierzy matrix, białek surowicy – albumin, fibrynogenu, transferyny, białek dopełniacza (5). Główne adhezyny grzybów stanowią białka ściany komórkowej: Als1-Als9 (ang. agglutinin-like sequence proteins), Hwp1 (ang. hyphae wall protein) – mannoproteina specyficzna dla formy strzępkowej C. albicans, białko Eap1p oraz Epa1p (ang. epithelial adhesion protein) – lecytyno-zależna adhezyna charakterystyczna dla komórek C. glabrata (4, 6). Wśród adhezyn grzybiczych są także białka niezwiązane ze ścianą komórkową Candida: Int1 (ang. integrin-like protein) – białko powierzchniowe blastospor i strzępek wiążące białka macierzy zewnątrzkomórkowej oraz białka błonowe Mnt1 i Pmt1. Najnowsze badania opisują nowy czynnik – kinazę Tor1, która może mieć bardzo szeroki wpływ regulacyjny na adhezyny. Kinaza ta znajduje się w szlaku regulującym zapotrzebowanie grzybów na składniki odżywcze. Ponadto białko to odgrywa dużą rolę w regulacji ekspresji genów specyficznych dla blastospor i formy micelialnej, włączając geny adhezyn – ALS1-3, HWP1, oraz inhibitorów filamentacji białek – Nrg1 i Tup1 (6). Dowiedziono, że białko Tor1 w odpowiedzi na warunki pokarmowe stymuluje do działania zarówno czynniki pobudzające, jak i inhibitory adhezji oraz filamentacji Candida spp.
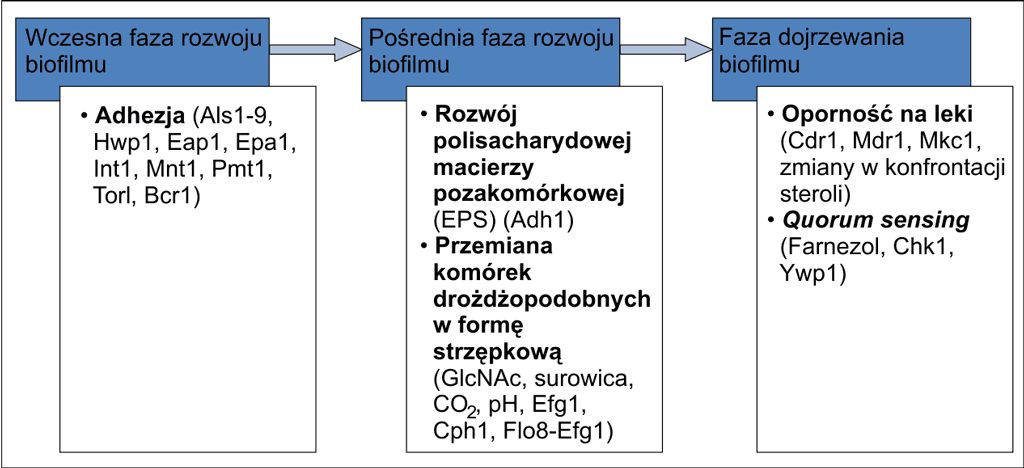
W zakażeniach odcewnikowych szczególnie istotne znaczenie w wirulencji szczepów Candida spp. ma zdolność tworzenia biofilmu. Biofilm Candida spp. to specyficzna struktura składająca się z komórek grzybów otoczonych zewnątrzkomórkowym śluzem, przylegających do powierzchni sztucznych, m.in. polimetakrylanu metylu, silikonu elastomerowego, polichlorku winylu (7, 8). Tworzenie się struktury biofilmu jest wielostopniowym i złożonym procesem, który opiera się głównie na adhezji komórek planktonicznych oraz wytworzeniu pozakomórkowej. Dojrzała forma biofilmu, w zależności od właściwości morfologicznych szczepu oraz warunków środowiska, składa się z komórek drożdżopodobnych i/lub strzępek (ryc. 2) (9-11). Do powstania dojrzałej złożonej formy biofilmu u C. albicans niezbędne są białko Efg1 oraz enzym N-acetyloglukozaminidaza – inicjujące tworzenie strzępek (4, 12). Wśród czynników etiologicznych fungemii odcewnikowych najwyższą zdolność do produkcji biofilmu posiadają szczepy z gatunków innych niż C. albicans, w szczególności gatunek C. parapsilosis. Grzyby tworzące strukturę biofilmu są oporne na wiele leków przeciwgrzybicznych. Lekooporność biofilmów Candida spp. jest związana z bardzo rozwiniętą komunikacją międzykomórkową odbywającą się wewnątrz tej struktury. Komunikacja ta zachodzi na drodze quorum sensing (3, 8). Obecnie trwają badania nad zastosowaniem inhibitorów quorum sensing jako substancji blokujących powstanie biofilmu. Alternatywą w terapii niszczącej strukturę biofilmu Candida spp. mogą być także inhibitory enzymów wpływających na transmisję β-1,3-glukanu do macierzy biofilmu. Glukany przeniesione ze ściany komórkowej do macierzy biofilmu mogą ograniczać penetrację leków do wnętrza tej struktury (13, 14). Zdolność budowania struktury biofilmu przez szczepy Candida spp. jest związana z aktywnością enzymatyczną grzybów. Do głównych, poznanych enzymów grzybów drożdżopodobnych zaliczamy: proteazy aspartylowe Sap (ang. secreted aspartyl protease), yapsinopodobne proteazy aspatylowe YPS (ang. yapsin-like protease), fosfolipazy, lipazy (Lip1-10), fosfomannoesterazę, N-acetyloglukozyloaminidazę (NAG) oraz α-glukozydazę (15, 16). Większość autorów w swoich pracach naukowych donosi o znaczącej roli proteaz w patogenezie zakażeń grzybiczych. Proteazy aspatylowe występują w komórkach C. albicans w postaci 10 izoenzymów. C. parapsilosis posiada proteazy kodowane przez geny SAP1-2-3, C. tropicalis geny SAP1-4, a C. dubliniensis – SAP1-4, SAP7-10 (15, 16). Izoenzymy Sap1-3 występują najczęściej w komórkach blastospor o zmniejszonej chorobotwórczości, najczęściej odpowiedzialnych jedynie za kolonizację nabłonka czy zakażenia błon śluzowych. Dowiedziono, że geny SAP ewoluowały i produkty ich ekspresji proteazy Sap1-3 mogą być wydzielane przez szczepy C. albicans także w zakażeniach systemowych. Proteazy Sap4-6 wytwarzane są przez formy strzępkowe o wysokiej zjadliwości, odpowiedzialne za kandydozy układowe (17). Prawdopodobnie proteazy Sap5-8 mają znaczenie w tworzeniu biofilmu. Produkty genu SAP7 mogą mieć szczególny wpływ na wirulencję szczepów wywołujących zakażenia wewnątrznaczyniowe. Szczepy C. glabrata posiadają geny YPS kodujące 11 proteaz yapsinopodobnych Yps1-11. Proteazy Yps odgrywają znaczącą rolę w utrzymywaniu integralności ściany komórkowej podczas zmiany fenotypowej szczepów C. glabrata (18).
Ryc. 2. Fazy rozwoju biofilmu C. albicans [wg (10), zmodyfikowano].
Cel PRACY
Celem badań retrospektywnych była ocena fenotypowych i genotypowych cech szczepów Candida spp. izolowanych od chorych z podejrzeniem fungemii odcewnikowych. Chorzy hospitalizowani byli w Centralnym Szpitalu Klinicznym w Warszawie przy ul. Banacha w latach 2005-2009.
Materiał i metody
Analizie poddano 90 szczepów Candida spp. pochodzących od 30 chorych (10 kobiet, 20 mężczyzn) hospitalizowanych w oddziałach CSK. Szczepy wyizolowano z 86 materiałów klinicznych, w tym 38 próbek krwi obwodowej, 22 próbek krwi z cewników naczyniowych oraz 26 fragmentów cewników naczyniowych (centralnych i obwodowych).
Hodowlę próbek krwi prowadzono na płynnych pożywkach transportowo-wrostowych w systemie BacT/Alert w temperaturze 37°C (bioMèrieux). Fragmenty cewników naczyniowych posiewano na podłoża wzrostowe metodą Maki oraz metodą Brun-Bruisson. Hodowlę grzybów prowadzono na podłożu Sabourauda w temperaturze 30°C. Przynależność gatunkową grzybów drożdżopodobnych oznaczano z zastosowaniem podłoża chromogennego CHROMagar Candida (bioMèrieux) oraz testu biochemicznego API ID32C (bioMèrieux) opartego o asymilację substratów węglowodanowych.
Powyżej zamieściliśmy fragment artykułu, do którego możesz uzyskać pełny dostęp.
Mam kod dostępu
- Aby uzyskać płatny dostęp do pełnej treści powyższego artykułu albo wszystkich artykułów (w zależności od wybranej opcji), należy wprowadzić kod.
- Wprowadzając kod, akceptują Państwo treść Regulaminu oraz potwierdzają zapoznanie się z nim.
- Aby kupić kod proszę skorzystać z jednej z poniższych opcji.
Opcja #1
29 zł
Wybieram
- dostęp do tego artykułu
- dostęp na 7 dni
uzyskany kod musi być wprowadzony na stronie artykułu, do którego został wykupiony
Opcja #2
69 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 30 dni
- najpopularniejsza opcja
Opcja #3
129 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 90 dni
- oszczędzasz 78 zł
Piśmiennictwo
1. Fernandez J, Erstad BL, Petty W, Nix DE: Time to positive culture and identification for Candida blood stream infections. Diagn Microbiol Infect Dis 2009; 64(4): 402-407.
2. Sulik-Tyszka B, Netsvyetayeva I, Jarzynka S et al.: Inwazyjne zakażenia grzybicze – nowe możliwości terapeutyczne. Zakażenia 2010; 10: 62-69.
3. Lim CS, Rosli R, Seow HF, Chong PP: Candida and invasive candidiasis: back to basics. Eur J Clin Microbiol Infect Dis 2012; 31(1): 21-31.
4. Huang G: Regulation of phenotypic transitions in the fungal pathogen Candida albicans. Virulence 2012; 3(3): 251-261.
5. Krutkiewicz A: Czynniki chorobotwórczości Candida albicans. Mikol Lek 2010; 17: 134-137.
6. Bastidas RJ, Heitman J, Cardenas ME: The protein kinase Tor1 regulates adhesin gene expression in Candida albicans. PLoS Pathog 2009; 5(2): e1000294.
7. Różalska B, Sadowska B: Zapobieganie powstawaniu biofilmów – perspektywiczne strategie ich eradykacji. Sepsis 2010; 3(2): 71-78.
8. Ramage G, Coco B, Sherry L et al.: In vitro Candida albicans biofilm induced proteinase activity and SAP8 expression correlates with in vivo denture stomatitis severity. Mycopathologia 2012; 174(1): 11-19.
9. Trofa D, Gacser A, Nosanchuk JD: Candida parapsilosis an Emerging Fungal Pathogen. Clin Microbiol Rev 2008; 21: 606-625.
10. Blankenship R, Mitchell A: How to build a biofilm: a fungal perspective. Current Opinion in Microbiology 2006; 9: 588-594.
11. Cuèllar-Cruz M, López-Romero E, Villagómez-Castro JC, Ruiz-Baca E: Candida species: new insights into biofilm formation. Future Microbiol 2012; 7(6): 755-771.
12. Yi S, Sahni N, Daniels KJ et al.: Alternative mating type configurations (a/α versus a/a or α/α) of Candida albicans result in alternative biofilms regulated by different pathways. PLoS Biol 2011; 9(8): e1001117.
13. Taff HT, Nett JE, Zarnowski R et al.: Candida biofilm-induced pathway for matrix glucan delivery: implications for drug resistance. PLoS Pathog 2012; 8(8): e1002848.
14. Shin JH, Kee S, Shin MG, Kim SH: Biofilm Production by Isolates of Candida Species Recovered from Nonneutropenic Patients: Comparison of Bloodstream Isolates with Isolates from Other Sources. J Clin Microbiol 2002; 40: 1244-1248.
15. Parra-Ortega B, Cruz-Torres H, Villa-Tanaca L, Hernández-Rodríguez C: Phylogeny and evolution of the aspartyl protease family from clinically relevant Candida species. Mem Inst Oswaldo Cruz 2009; 104(3): 505-512.
16. Tavanti A, Hensgens LA, Mogavero S et al.: Genotypic and phenotypic properties of Candida parapsilosis sensu stricto strains isolated from different geographic regions and body sites. BMC Microbiol 2010; 10: 203.
17. Gültekin B, Eyigör M, Tiryaki Y et al.: Investigation of antifungal susceptibilities and some virulence factors of Candida strains isolated from blood cultures and genotyping by RAPD-PCR. Mikrobiyol Bul 2011; 45(2): 306-317.
18. Ferrari S, Sanguinetti M, De Bernardis F et al.: Loss of mitochondrial functions associated with azole resistance in Candida glabrata results in enhanced virulence in mice. Antimicrob Agents Chemother 2011; 55(5): 1852-1860.
19. Kurnatowska AJ: Activity of hydrolytic enzymes of Candida albicans strains isolated from patients with periodontal and membrane mucosae of oral cavity diseases. Mycopathologia 1998; 141(2): 105-109.
20. Giri S, Kindo AJ: A review of Candida species causing blood stream infection. Indian J Med Microbiol 2012; 30: 270-278.
21. Almirante B, Rodríguez D, Cuenca-Estrella M et al.: Epidemiology, risk factors, and prognosis of Candida parapsilosis bloodstream infections: case-control population-based surveillance study of patients in Barcelona, Spain, from 2002 to 2003. J Clin Microbiol 2006; 44(5): 1681-1685.
22. Alvarez FJ, Konopka JB: Identification of an N-acetylglucosamine transporter that mediates hyphal induction in Candida albicans. Mol Biol Cell 2007; 18(3): 965-975.
23. Mohan das V, Ballal M: Proteinase and phospholipase activity as virulence factors in Candida species isolated from blood. Rev Iberoam Micol 2008; 25(4): 208-210.
24. Dąbkowska M: Badanie aktywności enzymatycznej szczepów Candida albicans izolowanych z materiałów klinicznych biorcy nerki. Mikol Lek, List do redakcji 2007; 14(1): 73-74.
25. Jayatilake JA, Samaranayake YH, Cheung LK, Samaranayake LP: Quantitative evaluation of tissue invasion by wild type, hyphal and SAP mutants of Candida albicans, and non-albicans Candida species in reconstituted human oral epithelium. J Oral Pathol Med 2006; 35(8): 484-491.
26. Kaur R, Ma B, Cormack BP: A family of glycosylphosphatidylinositol-linked aspartyl proteases is required for virulence of Candida glabrata. Proc Natl Acad Sci U S A 2007; 104(18): 7628-7633.