© Borgis - Postępy Fitoterapii 3/2004, s. 126-137
Justyna Makowska-Wąs, Zbigniew Janeczko
Biodostępność polifenoli roślinnych
Bioavailability of plant polyphenols
Katedra Farmakognozji Collegium Medicum Uniwersytetu Jagiellońskiego w Krakowie
Kierownik: prof. UJ dr hab. Zbigniew Janeczko
Summary
Flavonoids comprise the most common group of plant polyphenols. More than 4000 different flavonoids have been described so far. The six major classes of flavonoids include flavones (e.g. apigenin, luteolin, diosmetin), flavonols (e.g. kaempferol, quercetin, myricetin), flavanones (e.g. naringenin, hesperedin), flavanols (e.g. catechin, gallocatechin), anthocyanidins (e.g. pelargonidin, cyanidin) and isoflavones (e.g. daidzein, genistein). Flavonoids occur in food primarily as glycosides and polymers. The daily consumption of total flavonoids, mainly in vegetables, fruits and beverages is described as 1-2g. Flavonoids are poorly absorbed from the gut and are subjected to degradation by intestinal microorganism to various phenolic acids. Phenolic acids are a subclass of polyphenols. Their metabolism is very similar to flavonoids.

Flawonoidy należą do związków polifenolowych, szeroko rozpowszechnionych w świecie roślinnym. Występują w nadziemnych częściach roślin, niejednokrotnie nadając barwę kwiatom czy owocom w zakresie od żółtej (flawonoidy) do czerwonej i fioletowej (antocyjany). Dotychczas opisano budowę ponad 4 000 związków z tej grupy.
Struktura flawonoidów oparta jest na układzie flawanu, składającego się z trzech pierścieni A, B i C (ryc. 1). Modyfikacje w obrębie heterocyklicznego pierścienia C prowadzą do powstania różnych związków flawonoidowych, takich jak flawony (np. apigenina, luteolina, diosmetyna) (ryc. 2), flawonole (np. kemferol, kwercetyna, myrycetyna) (ryc. 3), flawanony (np. naryngenina, hesperydyna) (ryc. 4), flawanole (np. katechina, galokatechina) (ryc. 5), antocyjanidyny (np. pelargonidyna, cyjanidyna) (ryc. 6), czy izoflawony (np. daidzeina, genisteina) (ryc. 7) (70). Dodatkowa różnorodność tych związków wynika z możliwej obecności ugrupowań hydroksylowych, metoksylowych, izoprenylowych, cukrów prostych powiązanych wiązaniem O- lub C-glikozydowym, czy tworzeniem struktur dimerycznych (2, 66). Zalicza się tu także połączenia, takie jak: flawonolignany (pochodne flawanonolu – sylibina), czy estry katechiny (galusan epigalokatechiny) (32).
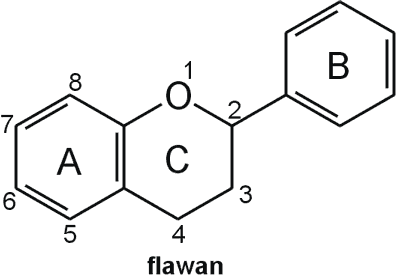
Ryc. 1. Wzór chemiczny flawanu.
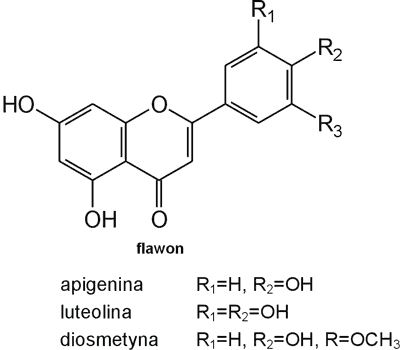
Ryc. 2. Pochodne flawonu.
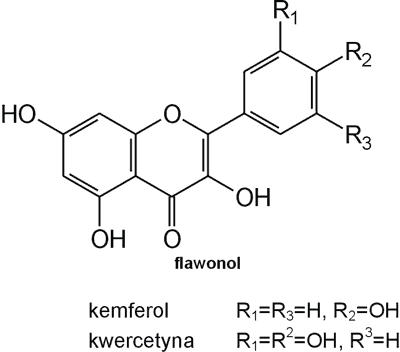
Ryc. 3. Pochodne flawonolu.
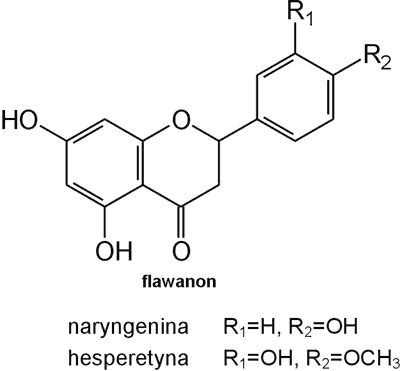
Ryc. 4. Pochodne flawanonu.
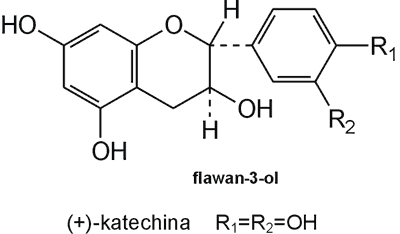
Ryc. 5. Pochodna flawanolu – (+)-katechina.
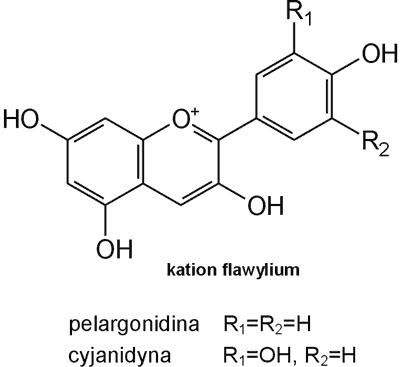
Ryc. 6. Pochodne antocyjanidyn.
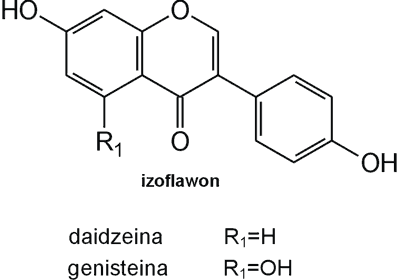
Ryc. 7. Pochodne izoflawonu.
Większość flawonoidów w roślinach występuje, obok wolnych aglikonów, w postaci połączeń glikozydowych (głównie O-glikozydy). W części cukrowej zazwyczaj występuje glukoza, także galaktoza, ramnoza, ksyloza, arabinoza. Cukry najczęściej przyłączone są do grupy hydroksylowej w pozycji C-7 we flawonach i izoflawonach; pozycji C-3 i C-7 w strukturach flawonoli oraz C-3 i C-5 w przypadku antocyjanidyn (66).
Związki flawonoidowe są składnikiem codziennej diety; źródła najczęściej spotykanych przedstawicieli poszczególnych klas przedstawiono w tabeli 1. Dzienne spożycie pochodnych flawonoidowych, głównie pod postacią warzyw, owoców i napojów, określa się na poziomie 1-2 g (30).
Już w latach 30. ubiegłego wieku laureat Nagrody Nobla Albert Szent-Györgyi odkrył wyraźną aktywność antyoksydacyjną flawonoidów (32). Późniejsze badania określiły mechanizmy warunkujące kierunki działania związków flawonoidowych: „wymiatanie” wolnych rodników, hamowanie peroksydacji lipidów, czy obniżenie aktywności enzymów. Stąd wynika ich zastosowanie jako środków ograniczających częstość występowania chorób serca, wątroby i pewnych typów nowotworów (32). Szeroko zakrojone badania epidemiologiczne dowiodły, że regularne spożywanie związków flawonoidowych może zmniejszać ryzyko śmiertelności z powodu chorób serca i układu krążenia (31, 40), a także występowania niektórych typów nowotworów (41). Polifenole, jako czynniki redukujące, wspólnie z innymi związkami o podobnej aktywności, jak witamina C, E i karoten, wykazują wpływ ochronny na tkanki wobec czynników stresu oksydacyjnego.
Związki o strukturze flawonoidowej obecne są w diecie, a także wchodzą w skład preparatów leczniczych stosowanych w chorobach serca i naczyń, zaburzeniach krążenia, głównie obwodowego (rutozyd, diosmina, hesperydyna), schorzeniach wątroby (sylimaryna), czy profilaktyki i łagodzenia objawów menopauzalnych (izoflawony sojowe). Należy więc, w kontekście ich działania, rozważyć stopień wchłaniania z przewodu pokarmowego człowieka oraz dalsze losy w ustroju.
W ciągu ostatnich lat pojawiło się wiele doniesień o wynikach badań nad absorpcją, metabolizmem i wydalaniem flawonoidów, prowadzonych w warunkach in vitro i in vivo. Wiele spornych kwestii zostało wyjaśnionych, jednakże, w związku z różnorodnością czynników wpływających na dostępność oraz kierunki metabolizmu jelitowego i wątrobowego, wyniki badań z różnych ośrodków nie zawsze pokrywają się. Pytanie podstawowe pozostaje nadal aktualne: czy absorbowane są aglikony, glikozydy, czy obie formy jednocześnie?
Aglikony flawonoidowe mają charakter hydrofobowy i mogą być transportowane przez błony biologiczne na drodze dyfuzji biernej (2). Połączenie z cukrem w postać glikozydową zmienia charakter związku na bardziej hydrofilny, co zmniejsza możliwość dyfuzji. b-Glukozydazy, obecne w aktywnym enzymatycznie nabłonku jelita cienkiego, umożliwiają wchłanianie wolnych aglikonów, poprzez rozszczepienie wiązania b-glikozydowego. Metabolizm flawonoidów zachodzi w wątrobie, przy udziale enzymów I fazy (hydroksylacja, demetylacja – cytochrom P 450) oraz II fazy (O-metylacja, sprzęganie z kwasem glukuronowym lub siarkowym), częściowo już w jelicie (32). Produkty metabolizmu związków flawonoidowych wydalane są z moczem oraz z żółcią, ulegając krążeniu jelitowo-wątrobowemu, co przedłuża czas eliminacji a także możliwego działania. Niewchłonięte oraz wydzielone z żółcią metabolity flawonoidów są przetwarzane przez mikroflorę jelitową, głównie w jelicie grubym. Enzymy bakteryjne mogą katalizować reakcje, takie jak hydroliza glukuronidów, siarczanów i glikozydów, dehydroksylacja, demetylacja, redukcja wiązania podwójnego, rozkład pierścienia C z utworzeniem fenolokwasów, a następnie ich dekarboksylację (76). Kwasy fenolowe mogą być absorbowane, ulegać koniugacji czy O-metylacji w wątrobie, a następnie wydaleniu z moczem (35). Absorpcja może mieć znaczenie dla całkowitej aktywności antyoksydacyjnej osocza, gdyż kwasy posiadające strukturę katecholową wykazują działanie wymiatające wolne rodniki (52).
Flawonole degradowane są do pochodnych kwasu fenylooctowego i fenylopropionowego. Rozerwanie pierścienia C katechin prowadzi do powstania fenylowalerolaktonów i kwasu hydroksyfenylopropionowego. Flawony i flawanony przekształcane są do kwasu fenylopropionowego, a następnie pochodnych kwasu benzoesowego (76).
FLAWONOLE
Do najczęściej spotykanych w przyrodzie związków flawonoidowych należą flawonole, a zwłaszcza kwercetyna i jej glikozydy. Znaleźć je można w cebuli, herbacie, czerwonym winie czy owocach, takich jak jabłka (tab. 1).
Tabela 1. Źródła polifenoli w pożywieniu (wg 2, 47, 66, 79).
Flawonole
kwercetyna
kemferol |
cebula, sałata, brokuły, ciemne winogrona, jabłka, herbata,
brokuły, grejpfrut, czarna herbata |
Flawony
luteolina
apigenina |
seler, cytryny, czerwona papryka
seler, pietruszka |
Flawanony
hesperetyna
naryngenina |
pomarańcze
grejpfruty |
Flawanole
(+)-katechina
(-)-epikatechina
proantocyjanidyny |
jabłka, winogrona, czerwone wino, herbata, czekolada
|
Izoflawony
genisteina
daidzeina |
soja, produkty sojowe, warzywa strączkowe
|
Antocyjany
cyjanidyna
delfinidyna |
owoce: czarnej porzeczki, truskawki, winorośli, wiśni, bzu czarnego,
owoce: aronii, borówki czernicy |
Fenolokwasy
kwas kawowy
kwas chlorogenowy
kwas ferulowy |
białe winogrona, oliwki, kapusta
jabłka, wiśnie, pomidory, brzoskwinie, gruszki,
zboża, pomidory, szparagi |
Działanie biologiczne kwercetyny wynika z jej wysokiej aktywności antyoksydacyjnej oraz możliwości hamowania enzymów, m.in. biorących udział w powstawaniu procesu zapalnego (65). Kwercetyna i jej pochodne, jak rutozyd (3-ramnoglukozyd kwercetyny), stosowane są w terapii wspomagającej niewydolność żylną kończyn dolnych oraz jako naturalne przeciwutleniacze.
Glikozydy flawonoidowe, podobnie jak hydroksykwasy, nie ulegają kwaśnej hydrolizie w żołądku (76). Zaproponowany mechanizm wchłaniania opiera się na aktywności endogennych b-glukozydaz obecnych w jelicie cienkim. Badania Day i wsp. (16) oraz Nemeth i wsp. (55) wykazały, że aktywne są szczególnie dwa enzymy: hydrolaza florydzynowa (EC 3.2.1.62) i b-glukozydaza cytozolowa (EC 3.2.1.21). Hydrolaza obecna jest w ścianie jelita cienkiego, na zewnętrznej powierzchni komórek nabłonka, gdzie odpowiada za hydrolizę laktozy. b-Glukozydaza cytozolowa występuje w narządach, takich jak wątroba, nerki i jelito cienkie u ssaków (15). Uważa się, że współuczestniczy ona w detoksykacji ksenobiotyków poprzez hydrolizę wiązania b-glukozydowego, co umożliwia następnie sprzęganie z kwasem glukuronowym i szybkie wydalanie z moczem lub żółcią. Powstałe po hydrolizie aglikony wchłaniane są przez ścianę jelita cienkiego na drodze dyfuzji biernej (76).
Przypuszcza się, że połączenia z glukozą mogą przechodzić przez błonę enterocytów podczas transportu aktywnego. Badania nad farmakokinetyką kwercetyny wykazały, że po podaniu doustnym jej glukozydy wchłaniane są w jelicie cienkim. Hollman i wsp. (36) określili absorpcję różnych form kwercetyny u zdrowych ochotników z przetoką kątniczą, co zapobiegało degradacji połączeń flawonoidowych przez mikroflorę jelitową. Wykazano, że absorpcja kwercetyny wynosiła 52±5% dla glukozydów obecnych w cebuli. Wolny aglikon i 3-O-rutynozyd kwercetyny wchłaniały się odpowiednio w ilości 24±9% i 17±15%.
Wchłanianie kwercetyny z cebuli jest do trzech razy większe niż z jabłek (48). Cebula zawiera głównie glukozydowe połączenia kwercetyny, natomiast w jabłkach kwercetyna występuje jako mieszanina glikozydów, takich jak galaktozydy, ramnozydy, arabinozydy, ksylozydy, glukozydy i rutynozydy (37, 48). Lepsza dostępność glukozydów kwercetyny, w porównaniu z aglikonem, wynika z udziału Na+-zależnego przenośnika glukozy (SGLT-1) w procesie absorpcji przez ścianę jelita (1, 2, 34). Transport aktywny 4´-O-glukozydu kwercetyny przy udziale SGLT-1 zademonstrowano na hodowli komórek nowotworu jelita grubego (Caco-2) (81).
W oparciu o wyniki dalszych badań stwierdzono, że dostępność 3-O-glukozydu i 4´-O-glukozydu kwercetyny jest bardzo zbliżona (56). U ludzi wchłanianie glukozydów zachodzi szybciej, a miejsce przyłączenia glukozy do aglikonu nie ma większego znaczenia dla absorpcji tych połączeń. (34, 56). Rutozyd ulega przekształceniu do 3-O-glukozydu kwercetyny przez enzymatyczne odszczepienie cząsteczki ramnozy (b-L-ramnozydaza), co znacznie zwiększa jego dostępność. Jednak proces ten zachodzi dopiero w okrężnicy pod wpływem mikroflory (56). Po podaniu ochotnikom cebuli, jabłek i rutozydu, kwercetyna pojawiła się w osoczu krwi odpowiednio po 0,7, 2,5 i 9 godzinach od momentu spożycia (37). Wyniki te potwierdzają udział bakterii jelitowych w procesie absorpcji rutozydu.
W wątrobie grupy hydroksylowe w pierścieniu B kwercetyny i jej glukozydów ulegają metylacji, co prowadzi do powstania m.in. izoramnetyny (3´-O-metylokwercetyny). W surowicy krwi, podobnie jak w moczu, nie stwierdzono wolnego aglikonu (kwercetyny) (76, 78). Głównymi metabolitami występującymi w moczu i żółci są glukuronidy kwercetyny, 3´-O-metylokwercetyny i 4´-O-metylokwercetyny (76). Po spożyciu cebuli, głównymi metabolitami oznaczanymi we krwi były 3´-siarczan i 3-glukuronid kwercetyny (17). Według Olthof´a i wsp. (56) tylko 3% przyjętej kwercetyny wydalane jest z moczem. Wskazuje to na znaczne nasilenie procesów metabolicznych w wątrobie oraz jelicie grubym (mikroflora).
Znaczna część przyjętej kwercetyny (niewchłonięta w jelicie cienkim oraz wydalona z żółcią) ulega przemianom pod wpływem flory jelitowej. Dochodzi do uwolnienia aglikonu z połączeń glikozydowych i glukuronianów, rozerwania pierścienia i powstania fenolokwasów, które następnie mogą być wchłaniane (56). Po degradacji połączeń kwercetyny przez mikroflorę powstają następujące kwasy: 3-hydroksycynamonowy, 3-hydroksyfenylooctowy, 3,4-dihydroksyfenylooctowy, 3-metoksy-4-hydroksyfenylooctowy (homowanilinowy) (28, 64, 76).
Rozerwanie heterocyklicznego pierścienia C powoduje zanik aktywności antyoksydacyjnej (73). Metabolity kwercetyny, takie jak 3´-O-metylokwercetyna, mogą nadal wykazywać działanie biologiczne; właściwości przeciwutleniające wykazano w badaniach in vitro (45). Produkty rozkładu rutozydu i kwercetyny: kwasy 3,4-dihydroksyfenylooctowy i 4-hydroksyfenylooctowy, wykazują silniejszą aktywność antyagregacyjną niż związki wyjściowe (75).
Całkowita eliminacja kwercetyny z organizmu zachodzi powoli; okres półtrwania wynosi około 25 godzin (36). Wynika to z łączenia się koniugatów z albuminami osocza, a także możliwości wchodzenia metabolitów do krążenia jelitowo-wątrobowego (72).
Jak przedstawiono powyżej, przekształcenie glikozydów kwercetyny w połączenia glukozydowe jest znaczącym krokiem do zwiększonego wchłaniania kwercetyny z pożywienia oraz preparatów leczniczych. Rutozyd, składnik wielu preparatów, może być nazywany „prolekiem”, gdyż dopiero jego metabolity ulegają wchłonięciu i wykazują aktywność biologiczną.
FLAWONY
Spośród flawonów w owocach i warzywach najczęściej występują glikozydy luteoliny i apigeniny (ryc. 2). W pożywieniu źródłem o największym znaczeniu są pietruszka i seler (tab. 1). W lecznictwie znalazła zastosowanie diosmina (7-rutynozyd diosmetyny), która wraz z flawanonem hesperydyną (7-rutynozyd hesperetyny), stosowana jest w terapii niewydolności krążenia żylnego i hemoroidów.
Przemiany metaboliczne flawonów są analogiczne do opisanych wcześniej dla kwercetyny. Połączenia glikozydowe są rozkładane w jelicie, wolne aglikony wchłaniają się z jelita lub ulegają degradacji pod wpływem enzymów bakteryjnych. 7-O-glukozyd luteoliny jest hydrolizowany do aglikonu przez endogenne b-glukozydazy, a następnie absorbowany w jelicie cienkim. Podczas przejścia przez śluzówkę jelita dochodzi do sprzęgania, głównie z kwasem glukuronowym (73). W osoczu luteolina występuje przede wszystkim w postaci monoglukuronianów (50).
W jelicie grubym mikroflora bakteryjna przetwarza flawony do pochodnych kwasu fenylopropionowego. W badaniach Booth i wsp. (7), po podaniu szczurom dożołądkowo diosminy w moczu oznaczali jako główny metabolit kwas 3-hydroksyfenylopropionowy. Po podaniu w analogiczny sposób diosmetyny wyniki były podobne, oznaczono dodatkowo glukuronid diosmetyny.
Flawony, takie jak diosmina i jej aglikon diosmetyna, mogą modyfikować metabolizm ksenobiotyków poprzez wpływ na enzymy cytochromu P 450, oba związki powodują wzrost aktywności CYP 1A1 (13, 32).
Na wchłanianie flawonoidów z preparatów leczniczych może mieć wpływ formulacja postaci leku. Mikronizacja diosminy znacząco polepsza wchłanianie tego związku, co potwierdziły badania Garnera i wsp. (27). Po podaniu zdrowym ochotnikom znakowanej 14C diosminy jej absorpcja z przewodu pokarmowego, mierzona całkowitą radioaktywnością związków wydalonych z moczem, wynosiła 57,9±20,2% dla zmikronizowanego i 32,7±18,8% dla niezmikronizowanego związku (27).
FLAWANONY
Flawanony są grupą związków, w które obfitują owoce cytrusowe, m.in. grejpfrut ( Citrus paradisi) i pomarańcza ( Citrus chinensis). Należą tu takie aglikony jak naryngenina, hesperetyna. 7-ramnoglukozyd naryngeniny (naryngina) znajduje się w soku, kwiatach i skórce grejpfruta, stanowiąc nawet 10% suchej masy (84). Hesperetyna i jej glikozydy to główne flawanony cytryn i słodkich pomarańczy. Szklanka soku pomarańczowego może zawierać 25-80 mg flawanonów w przeliczeniu na aglikon (46).
Naryngenina i hesperetyna mogą być wchłaniane z pożywienia, co potwierdzono oznaczając te związki w ludzkim osoczu oraz moczu, odpowiednio po podaniu soku grejpfrutowego i pomarańczowego oraz czystych związków (84). W przewodzie pokarmowym dochodzi do rozkładu połączeń glikozydowych; wchłaniane mogą być aglikony. Oznaczono szczepy bakterii odpowiedzialne za rozkład narynginy do naryngeniny oraz hesperydyny do wolnej hesperetyny; w zależności od składu mikroflory obserwowano dużą osobniczą rozbieżność w absorpcji tych związków (84).
Znaczna część wchłoniętej naryngeniny pojawia się w moczu w postaci połączeń z kwasem glukuronowym, a koniugacja zachodzi w wątrobie lub już w jelicie cienkim z pomocą UDP-glukuronylotransferazy. Badania na zdrowych ochotnikach wykazały, że w moczu 13% flawanonów było mieszanymi połączeniami z kwasem siarkowym i glukuronowym (46, 88). Wyniki badań in vitro prowadzonych na izolowanym jelicie cienkim, bądź jego homogenatach sugerują, że monoglikozydy mogą być rozkładane w jelicie cienkim, diglikozydy natomiast przechodzą do jelita grubego (12, 15). Po 24 godzinach od spożycia soku pomarańczowego eliminacja z moczem metabolitów wynosiła blisko 98% (46).
W badaniach na szczurach, stosując znakowaną 14C hesperetynę wykazano, że 40% wchłoniętego związku dociera do wątroby (84). Tam przez hydroksylację naryngeniny i demetylację hesperetyny, powstaje eriodiktiol, główny metabolit obu tych związków (84). Hydroksylacja naryngeniny zachodzi przy udziale wątrobowego izoenzymu cytochromu P 450 (CYP 1A), natomiast hesperetyna jest substratem dla CYP 1A1 i CYP 1B1 prowadzących demetylację (32). Flawanony mogą być specyficznymi induktorami cytochromów (np. CYP 2B1/2), jednak naryngenina wykazuje działanie hamujące wobec CYP 3A4 w enterocytach jelita cienkiego. Wraz z bergamotyną, furanokumaryną występującą także w Citrus paradisi, naryngenina odpowiada za upośledzenie metabolizmu leków podawanych razem z sokiem grejpfrutowym (14, 22, 46). Częstość występowania interakcji soku z grejpfruta z innymi lekami może być uzależniona od indywidualnych różnic w składzie mikroflory jelitowej, która powoduje rozkład glikozydów i uwolnienie łatwiej wchłanialnych aglikonów.
U szczurów główną drogą wydalania flawonoidów jest wydzielanie do żółci. Flawanony są wchłaniane zwrotnie i wchodzą do krążenia jelitowo-wątrobowego lub ulegają rozkładowi pod wpływem enzymów bakteryjnych w okrężnicy (84). Powstające hydroksykwasy mogą następnie być wchłaniane. Po podaniu szczurom naryngeniny w moczu oznaczono kwas 4-hydroksyfenylopropionowy (7), kwas 4-hydroksycynamonowy i 4-hydroksybenzoesowy (46). Niektórzy autorzy sądzą, że jedynie kwas 4-hydroksyfenylopropionowy jest produktem rozkładu bakteryjnego, a pozostałe metabolity powstają w wątrobie (84).
Przygotowanie odwarów z naowocni Citrus grandis z dodatkiem miodu powodowało zwiększenie stężenia narynginy w wyciągu. Lecz jednocześnie po podaniu zdrowym ochotnikom zmniejszyło wchłanianie tego związku, prawdopodobnie na skutek zahamowania powstawania aglikonu (naryngeniny) (39). Wyniki te wskazują na wpływ innych składników pożywienia na dostępność flawonoidów.
POCHODNE FLAWAN-3-OLU
Herbata, czyli suszone i odpowiednio preparowane liście Camelia sinensis, zawiera wiele związków, jednak jej aktywność biologiczna, taka jak obniżanie poziomu lipidów, działanie przeciwzapalne, przeciwbakteryjne, antymutagenne i antyoksydacyjne, wynika głównie z zawartości frakcji polifenoli, z największym udziałem katechin (91). W herbacie znajdują się m.in. wolne aglikony, takie jak (–)-epikatechina (EC) i (+)-katechina oraz estry kwasu galusowego: galusan 3-(–)-epikatechiny (ECG), galusan 3-(–)-epigalokatechiny (EGCG). Związki te występują także w czekoladzie (głównie (+)-katechina i (–)-epikatechina), czy czerwonym winie (m.in. (+)-katechina) (4, 6, 20).
Flawanole, w odróżnieniu od pozostałych pochodnych flawonoidowych, w roślinach występują jako formy niezglikozydowane. W związku z tym aktywność b-glukozydazy nie ma wpływu na proces wchłaniania. Stosunkowo dobrze absorbowana jest (+)-katechina, jednak inne pochodne flawonoidowe herbaty charakteryzują się niską absorpcją i dostępnością u ludzi (82). Udowodniono wpływ śliny na estry katechiny zawarte w zielonej herbacie; odłączenie reszty kwasu galusowego z galusanu epigalokatechiny zaobserwowali Yang i wsp. (87). Podstawniki hydroksylowe ugrupowań galoilowych mogą wiązać się z białkami powierzchniowymi błon oraz obecnymi w ślinie, co upośledza wchłanianie przez obniżenie rozpuszczalności (91). Podczas pasażu pokarmu z żołądka do jelita cienkiego może dochodzić do oksydacji galusanu epigalokatechiny z wytworzeniem produktów dimerycznych, które mają wyższą aktywność neutralizacji wolnych rodników i wiązania jonów żelaza niż EGCG (75).
Flawan-3-ole w jelicie cienkim ulegają sprzęganiu z kwasem glukuronowym, tworząc bardziej polarne koniugaty 5- lub 7-O-glukuronidowe oraz 3´- lub 4´-O-metyloglukuronidy (63). W innych procesach, prowadzonych przez metylotransferazy, powstają 3´-O-metylowe pochodne; ich aktywność antyoksydacyjna jest zmniejszona (20, 75). Glukuronidy katechiny i epikatechiny wykazują podobną do wolnych związków aktywność wymiatania wolnych rodników (38).
Badania prowadzone na myszach i szczurach potwierdziły wchłanianie z jelita galusanu epigalokatechiny (44, 53). EGCG oznaczany w osoczu w 50-90% był związany z kwasem glukuronowym. Badania Zhu i wsp. (91) wykazały największe stężenie katechin we krwi w 2 godziny po podaniu szczurom frakcji zawierającej EC, ECG i EGCG. Związki te w znacznym stopniu przenikają do tkanek, co mimo niskiej dostępności biologicznej, warunkuje ich korzystne działanie. Po podaniu dożylnym nieskoniugowany galusan epigalokatechiny oznaczano m.in. w płucach i prostacie (44).
Znaczne modyfikacje struktury zachodzą w okrężnicy, gdzie bakterie jelitowe degradują flawanole do kwasów fenolowych. Niektóre z nich mogą być wchłaniane i wywierać własne działanie biologiczne. Degradacja w okrężnicy prowadzi do rozkładu układu flawanu i wytworzenia prostszych związków fenolowych, jak walerolaktony i kwas hydroksyfenylopropionowy (63).
Jak wynika z badań Roodenburga i wsp. (69) spożywanie zielonej lub czarnej herbaty powoduje wzrost stężenia katechin we krwi, z towarzyszącym temu procesowi wzrostem aktywności antyoksydacyjnej, mierzonej zdolnością aktywności osocza do redukcji jonów żelaza (FRAP). Mleko, podawane z czarną herbatą, nie hamowało wchłaniania katechin, ani nie znosiło obserwowanego wzrostu aktywności przeciwutleniającej (69). Polepszenie absorpcji flawonoidów można uzyskać podając je wraz z substancjami emulgującymi. Podanie katechiny wraz z fosfolipidami zwiększyło jej wchłanianie (61).
Proantocyjanidyny są związkami polimerycznymi, których podstawowa struktura składa się z jednostek polihydroksyflawan-3-olu połączonych wiązaniami C-C (26). Proantocyjanidyny mają wybitną aktywność wymiatania wolnych rodników i hamowania oksydacji LDL in vitro. Właściwości te potwierdzono także badając wpływ na żywy organizm (szczury) (42). Oligomery proantocyjanidynowe uważane są w powszechnej opinii za najskuteczniejsze przeciwutleniacze roślinne. Szczególnie dużo związków tego typu znajduje się w czekoladzie, jabłkach, żurawinie czy czerwonym winie (tab. 1) (29). Wysoką ich zawartością charakteryzują się owoce winorośli; wyciągi z nasion stosowane są jako środek wspomagający leczenie przewlekłej niewydolności żylnej i mikroangiopatii (26).
Inkubacja oligomerów procyjanidyn (dimery – heksamery) w środowisku ludzkiej śliny przez 30 minut nie powodowała zmian w budowie tych związków, a także nie zaobserwowano rozkładu większych cząsteczek na mniejsze jednostki (75). Stwierdzono natomiast, że dimery proantocyjanidynowe mają wysokie powinowactwo do wiązania się z białkami śliny bogatymi w prolinę. Szczególnie wysoką aktywność wykazują estry galoilowe epikatechiny, a także galusany dimerów procyjanidyny typu B2 (25).
Według Spencer´a i wsp. (74) oligomery procyjanidynowe (od trimerów do dekamerów) izolowane z Theobroma cacao są niestabilne w kontakcie z sokiem żołądkowym ex vivo i tworzą monomery, dimery epikatechiny oraz inne jednostki oligomeryczne. Z przeprowadzonych przez Rios i wsp. (67) doświadczeń na zdrowych ochotnikach wynika, że w środowisku soku żołądkowego, po podaniu napoju z kakaowca, związki oligomeryczne pozostają niezmienione.
Oligomery procyjanidynowe o dużej masie cząsteczkowej odznaczają się wysokim powinowactwem do struktur białkowych ściany jelita (70). W eksperymentach z użyciem jednowarstwowej hodowli komórek nowotworu jelita grubego (Caco-2) wykazano, że jedynie dimery i trimery są w stanie przenikać przez nabłonek jelita (19). Oznaczono dimer procyjanidynowy B2, (–)-epikatechinę i (+)-katechinę we krwi u ludzi, już 30 minut po podaniu wyciągu z nasion kakaowca (38).
Powyżej zamieściliśmy fragment artykułu, do którego możesz uzyskać pełny dostęp.
Mam kod dostępu
- Aby uzyskać płatny dostęp do pełnej treści powyższego artykułu albo wszystkich artykułów (w zależności od wybranej opcji), należy wprowadzić kod.
- Wprowadzając kod, akceptują Państwo treść Regulaminu oraz potwierdzają zapoznanie się z nim.
- Aby kupić kod proszę skorzystać z jednej z poniższych opcji.
Opcja #1
29 zł
Wybieram
- dostęp do tego artykułu
- dostęp na 7 dni
uzyskany kod musi być wprowadzony na stronie artykułu, do którego został wykupiony
Opcja #2
69 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 30 dni
- najpopularniejsza opcja
Opcja #3
129 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 90 dni
- oszczędzasz 78 zł
Piśmiennictwo
1. Ader P., et al.: Interaction of quercetin glucosides with the intestinal sodium/glucose co-transporter (SGLT-1). Cancer Lett. 2000, 162, 175. 2.Aherne S.A., O´Brien N.M.: Dietary flavonols: chemistry, food content, and metabolism. Nutrition 2002, 18, 75. 3.Arcari M., et al.: A new inclusion complex of silibin and beta-cyclodextrins: in vitro dissolution kinetics and in vivo absorption in comparison with traditional formulations. Biol. Pharm. Bull. 1992, 131, 205. 4.Arts I.C.W., et al.: Chocolate as a source of tea flavonoids. Lancet 1999, 354, 488. 5.Bhattaram V.A., et al.: Pharmacokinetics and bioavailability of herbal medicinal products. Phytomed. 2002, 9, 1. 6. Bombardelli E., Morazzoni P.: Vitis vinifera L. Fitoterapia 1995, 66, 291. 7.Booth A.N., et al.: Metabolic fate of hesperidin, eriodictyol, homoeriodictyol, and diosmin. J. Biol. Chem. 1958, 230, 661. 8.Breinholt V.M., et al.: In vitro investigation of cytochrome P 450-mediated metabolism of dietary flavonoids. Food Chem. Toxicol. 2002, 40, 609. 9.Chang Y.C., Nair M.G.: Metabolites of daidzein and genistein and their biological activities. J. Nat. Prod. 1995, 58, 1901. 10.Chang Y.C., Nair M.G.: Metabolism of daidzein and genistein by intestinal bacteria. J. Nat. Prod. 1995, 58, 1892. 11.Chen C.Y., et al.: High-genistin isoflavone supplementation modulated erythrocyte antioxidant enzymes and increased running endurance in rats undergoing one session of exhausting exercise – a pilot study. Pakist. J. Nutr. 2002, 1, 1. 12.Choudhury R., et al.: Interactions of the flavonoid naringenin in the gastrointestinal tract and the influence of glycosylation. Biochem. Biophys. Res. Comm. 1999, 265, 410. 13.Ciolino H.P., et al.: Diosmin and diosmetin are agonists of the aryl hydrocarbon receptor that differentially affect cytochrome P 540 1A1 activity. Cancer Res. 1998, 58, 2754. 14.Dahan A., Altman H.: Food-drug interaction: grapefruit juice augments drug bioavailability – mechanism, extend and relevance. Eur. J. Clin. Nutr. 2004, 58, 1. 15.Day A.J., et al.: Deglycosylation of flavonoid and isoflavonoid glycosides by human small intestine and liver b-glucosidase activity. FEBS Lett. 1998, 436, 71. 16.Day A.J., et al.: Dietary flavonoid and isoflavone glycosides are hydrolysed by the lactase site of lactase phlorizin hydrolase. FEBS Lett. 2000, 468, 166. 17.Day A.J., et al.: Human metabolism of dietary flavonoids: identification of plasma metabolites of quercetin. Free Rad. Res. 2001, 35, 941. 18.Deprez S., et al.: Polymeric proanthocyanidins are catabolized by human colonic microflora into low-molecular-weight phenolic acids. J. Nutr. 2000, 130, 2733. 19.Deprez S., et al.: Transport of proanthocyanidin dimer, trimer and polymer across monolayers of human intestinal epithelial Caco-2 cells. Antiox. Redox. Signal 2001, 3, 957. 20.Donovan J.L., et al.: Catechin is present as metabolites in human plasma after consumption of red wine. J. Nutr. 1999, 129, 1662. 21. Ebadi M.: Pharmacodynamic basis of herbal medicine. CRC Press 2002. 22.Erlund I.: Chemical analysis and pharmacokinetics of the flavonoids quercetin, hesperetin and naringenin in humans. Academic dissertation. Helsinki 2002. 23.Felgines C., et al.: Strawberry anthocyanins are recovered in urine as glucuro- and sulphoconjugates in humans. J. Nutr. 2003, 133, 1296. 24.Frank T., et al.: Bioavailability of anthocyanidin-3-glucosides following consumption of red grape juice. Can. J. Physiol. Pharmacol. 2003, 81, 423. 25. Freitas V. de, Mateus N.: Structural features of procyanidin interaction with salivary proteins. J. Agric. Food Chem. 2001, 49, 940. 26.Gabetta B., et al.: Characterisation of proanthocyanidins from grape seeds. Fitoterapia 2000, 71, 162. 27.Garner R.C., et al.: Comparison of the absorption of micronized (Daflon 500 mg) and non micronized 14C-diosmin tablets after oral administration to healthy volunteers by accelerator mass spectrometry and liquid scintillation counting. J. Pharm. Sci. 2002, 91, 32. 28.Gräfe U., Veit M.: Urinary metabolites of flavonoids and hydroxycinnamic acids in humans after application of a crude extract from Equisetum arvense L. Phytomed. 1999, 6, 239. 29.Hammerstone J.F., et al.: Procyanidin content and variation in some commonly consumed foods. J. Nutr. 2000, 130, 2086. 30.Havsteen B.H.: The biochemistry and medical significance of the flavonoids. Pharmacol. Ther. 2002, 96, 67. 31.Hertog M.G.L., et al.: Dietary antioxidant flavonoids and risk of coronary heart disease: the Zuphten Elderly Study. Lancet 1993, 342, 1007. 32.Hodek P., et al.: Flavonoids-potent and versatile biologically active compounds interacting with cytochromes P 450. Chem. Biol. Interact. 2002, 139, 1. 33. Hof K.H. van het, et al.: Bioavailability of catechins from tea: the effect of milk. Eur. J. Clin. Nutr. 1998, 52, 356. 34. Hollman P.C.H., et al.: Absorption of dietary quercetin glycosides and quercetin in healthy ileostomy volunteers. Am. J. Clin. Nutr. 1995, 62, 1276. 35.Hollman P.C.H., Katan M.B.: Absorption, metabolism and health effects of dietary flavonoids in man. Biomed. Pharmacother. 1997, 51, 305. 36.Hollman P.C.H., et al.: Bioavailability of the dietary antioxidant flavonol quercetin in man. Cancer Lett. 1997, 114, 139. 37.Hollman P.C.H., et al.: Relative bioavailability of the antioxidant flavonoid quercetin from various foods in man. FEBS Lett. 1997, 418, 152. 38.Holt R.R., et al.: Procyanidin dimer B2 (epicatechin-4b-epicatechin) in human plasma after the consumption of a flavanol-rich cocoa. Am. J. Clin. Nutr. 2002, 76, 798. 39.Hou Y.C., et al.: Effect of honey on naringin absorption from a decoction of the pericarps of Citrus grandis. Planta Med. 2000, 66, 439. 40.Huxley R.R., Neil H.A.W.: The relation between dietary intake and coronary heart disease mortality: a meta-analysis of prospective cohort studies. Eur. J. Clin. Nutr. 2003, 57, 904. 41.Knekt P., et al.: Flavonoid intake and risk of chronic diseases. Am. J. Clin. Nutr. 2002, 76, 560. 42.Koga T., et al.: Increase of antioxidative potential of rat plasma by oral administration of proanthocyanidin-rich extract from grape seeds. J. Agric. Food Chem. 1999, 47, 1892. 43.Kong J.M., et al.: Analysis and biological activities of anthocyanins. Phytochem. 2003, 64, 923. 44. Lambert J.D., et al.: Epigalocatechin-3-gallate is absorbed but extensively glucuronidated following oral administration to mice. J. Nutr. 2003, 133, 4172. 45.Manach C., et al.: Quercetin is recovered in human plasma as conjugated derivatives which retain antioxidant properties. FEBS Lett. 1998, 426, 331. 46. Manach C., et al.: Bioavailability in humans of the flavanones hesperidin and narirutin after the ingestion of two doses of orange juice. Eur. J. Clin. Nutr. 2003, 57, 235. 47.Manach C., et al.: Polyphenols: food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727. 48.McAnlis G.T., et al.: Absorption and antioxidant effects of quercetin from onions, in man. Eur. J. Clin. Nutr. 1999, 53, 92. 49.Messina M.: A healthy look at sweet cherries. www.nwcherries.com. 50.Miyake Y., et al.: Identification and antioxidant activity of flavonoid metabolites in plasma and urine of eriocitrin-treated rats. J. Agric. Food Chem. 2000, 48, 3217. 51.Morazzoni P., et al.: Comparative pharmacokinetics of silipide and silimarin in rats. Eur. J. Drug Metab. Pharmacokinet. 1993, 18, 289. 52.Murkovic M., et al.: Analysis of anthocyanins in plasma for determination of their bioavailability. J. Food Comp. Analys. 2000, 13, 291. 53.Nakagawa K., Miyazawa T.: Absorption and distribution of tea catechin, (–)-epigallocatechin-3-gallate, in the rat. J. Nutr. Sci. Vitaminol 1997, 43, 679. 54.Narayan M.S., et al.: Antioxydant effect of anthocyanin on enzymatic and non-enzymatic lipid peroxidation. Prostagl. Leukotr. Essent. Fatty Acids 1999, 60, 1. 55.Nemeth K., et al.: Deglycosylation by small intestinal epithelial cell beta-glucosidases is a critical step in the absorption and metabolism of dietary flavonoid glycosides in humans. Eur. J. Nutr. 2003, 42, 29. 56.Olthof M.R., et al.: Bioavailabilities of quercetin-3-glucoside and quercetin-4´-glucoside do not differ in humans. J. Nutr. 2000, 130, 1200. 57.Olthof M.R., et al.: Chlorogenic acid, quercetin-3-rutinoside and black tea polyphenols are extensively metabolized in humans. J. Nutr. 2003, 133, 1806. 58.Passamonti S., et al.: The interaction of anthocyanins with bilitranslocase. Biochem. Biophys. Res. Commun. 2002, 296, 631. 59.Passamonti S., et al.: The stomach as a site for anthocyanins absorption from food. FEBS Lett. 2003, 544, 210. 60.Pietta P.G.: Flavonoids as antioxidants. J. Nat. Prod. 2000, 63, 1035. 61. Piscuła M.K., Terao J.: Quercetin´s solubility affects its accumulation in rat plasma after oral administration. J. Agric. Food Chem. 1998, 46, 4313. 62.Prior R.L.: Fruits and vegetables in the prevention of cellular oxidative damage. Am. J. Clin. Nutr. 2003, 78, 570. 63.Rechner A.R., et al.: The metabolic fate of dietary polyphenols in humans. Free Rad. Biol. Med. 2002, 33, 220. 64.Rechner A.R., et al.: Colonic metabolism of dietary polyphenols: influence of structure on microbial fermentation products. Free Rad. Biol. Med. 2004, 36, 212. 65.Rice-Evans C.A., et al.: Structure-antioxidant activity relationship of flavonoids and phenolic acids. Free Rad. Biol. Med. 1996, 20, 933. 66.Rice-Evans C.A., et al.: Antioxidant properties of phenolic compounds. Trends in plant science 1997, 2, 152. 67.Rios L.Y., et al.: Cocoa procyanidins are stable during gastric transit in humans. Am. J. Clin. Nutr. 2002, 76, 1106. 68.Rohdewald P.: Bioavailability and metabolism of Pycnogenol(r). Eur. Bull. Drug Res. 1999, 7, 2. 69.Roodenburg A.J.C., et al.: Bioavailability and antioxidant activity of tea flavonoids. 2-nd International Electronic Conference on Synthetic Organic Chemistry (ECSOC-2), September 1998. www.mdpi.org. 70.Ross J.A., Kasum C.M.: Dietary flavonoids: bioavailability, metabolic effects, and safety. Ann. Rev. Nutr. 2002, 22, 19. 71.Schandalik R., et al.: Pharmacokinetics of silybin in bile following administration of silipide and silymarin in cholecystectomy patients. Arzneim.-Forsch. 1992, 42, 964. 72.Skibola C.F., Smith M.T.: Potential health impacts of excessive flavonoid intake. Free Rad. Biol. Med. 2000, 29, 375. 73.Spencer J.P.E., et al.: The small intestine can both absorb and glucuronidate luminal flavonoids. FEBS Lett. 1999, 458, 224. 74.Spencer J.P.E., et al.: Decomposition of cocoa procyaniodins in the gastric milieu. Biochem. Biophys. Res. Commun. 2000, 272, 236. 75.Spencer J.P.E.: Metabolism of tea flavonoids in the gastrointestinal tract. J. Nutr. 2003, 133, 3255. 76.Stahl W., Vet al.: Bioavailability and metabolism. Mol. Aspects Med. 2002, 23, 39. 77.Strack D., Wray V.: Anthocyanins. In: Methods in plant biochemistry (P.M. Dey, J.B. Harborne). Vol. 1. Plant phenolics. Academic Press 1989. 78.Texier O., et al.: Comparison of the bioavailability of dietary quercetin and catechin in rats. 2-nd International Electronic Conference on Synthetic Organic Chemistry (ECSOC-2), September 1998. www.mdpi.org. 79.USDA database for the flavonoid content of selected foods – 2003. http://www.nal.usda.gov/fnic/foodcomp/data/flav.html. 80.Virgilli F., et al.: Ferulic acid excretion as a marker of consumption of a French Maritime Pine ( Pinus maritima) bark extract. Free Rad. Biol. Med. 2000, 28, 1249. 81.Walgren R.A., et al.: Cellular uptake of dietary flavonoid quercetin-4´-b-glucoside by sodium-dependent glucose transporter SGLT1. J. Pharmacol. Exp. Ther. 2000, 294, 837. 82.Walle T.: Absorption and metabolism of flavonoids. Free. Rad. Biol. Med. 2004, 36, 829. 83.WHO Monograph on selected medicinal plants. Vol. 1. Geneva 1999. 84.Wilcox L.J., et al.: Antiatherogenic properties of naringenin, a citrus flavonoid. Cardiovasc. Drug Rev. 1999, 17, 160. 85.Williams C.A., Harborne J.B.: Isoflavonoids. In: Methods in plant biochemistry (P.M. Dey, J.B. Harborne). Vol. 1. Plant Phenolics. Academic Press 1989. 86.Wu X., et al.: Absorption and metabolism of anthocyanins in elderly woman after consumption of elderberry or blueberry. J. Nutr. 2002, 132, 1865. 87.Yang C.S., et al.: Human salivary tea catechin levels and catechin esterase activities: implication in human cancer prevention studies. Cancer Epidemiol. Biomarkers Prev. 1999, 8, 83. 88.Yang C.Y., et al.: Determination of hesperetin and its conjugate metabolites in serum and urine. J. Food Drug Anal. 2002, 10, 143. 89. Yamamoto S., et al.: Soy, isoflavones and breast cancer risk in Japan. J. Nat. Cancer Inst. 1995, 12, 906. 90. Yasuda T., et al.: Urinary and biliary metabolites of daidzin and daidzein in rats. Biol. Pharm. Bull. 1994, 17, 1369. 91.Zhu M., et al.: Oral absorption and bioavailability of tea catechins. Planta Med. 2000, 66, 444.
