© Borgis - Postępy Nauk Medycznych 1/2010, s. 29-34
Andrzej Cieśla1, *Tomasz Mach1, Krystyna Pierzchała-Koziec2, Joanna Zubel2, Paweł Skwara1
Czynnik transformujący β1 wskaźnikiem progresji przewlekłego wirusowego zapalenia wątroby
Transforming growth factor β1 as an indicator of the progression of chronic viral hepatitis
1Katedra Gastroenetrologii, Hepatologii i Chorób Zakaźnych, Uniwersytet Jagielloński Collegium Medicum w Krakowie
Kierownik Katedry: prof. dr hab. med. Tomasz Mach
2Katedra Fizjologii i Endokrynologii Zwierząt Uniwersytetu Rolniczego w Krakowie
Kierownik Katedry: prof. dr hab. Krystyna Pierzchała-Koziec
Streszczenie
Czynnik transformujący β1 (TGF β1) jest główną cytokiną warunkującą postęp włóknienia w przewlekłym uszkodzeniu wątroby. Mechanizmy określające szybkość progresji przewlekłego zapalenia wątroby o etiologii wirusowej (pwzw) nie są w pełni poznane.
Celem badania była identyfikacja czynników warunkujących postęp włóknienia w pwzw.
W grupie 78 chorych, w tym 52 z pwzw C, 19 z pwzw B i 7 z niealkoholową chorobą stłuszczeniową wątroby oraz autoimmunologicznym zapaleniem wątroby oznaczano stężenie wątrobowe TGF β1 (Quantikine Immunoassay Kit). Uzyskane wyniki porównano w grupach zróżnicowanych pod względem zaawansowania procesu zapalno-martwiczego i włóknienia ocenianego badaniem histologicznym bioptatu wątroby (wg skali Batts'a i Ludwiga), insulinooporności (HOMA-IR>2,259), aktywności w surowicy krwi aminotransferazy alaninowej, asparaginianowej, γ-glutamylotranspeptydazy (γ-GTP), wiremii HCV i HBV, osiągnięcia trwałej odpowiedzi wirusologicznej (SVR) leczonych z pzw C interferonem α, wskaźnika masy ciała (BMI), masy tkanki tłuszczowej oraz obecności zespołu metabolicznego (EGIR 1999).
TGF β1 korelował z aktywnością procesu zapalno-martwiczego ocenianego histologicznie (p<0,00001) i aktywnością AlAT (p=0,0177), a także zaawansowaniem włóknienia (p=0,0002). Wyższe stężenie wątrobowego TGF β1 było obserwowane u chorych ze stłuszczeniem wątroby (p=0,0144), zwiększoną aktywnością γ-GTP (p=0,0296) oraz z insulinoopornością (p=0,057). W zakresie pozostałych badanych zmiennych nie stwierdzono statystycznie istotnego zróżnicowania TGF β1.
Wyniki pracy potwierdziły ścisły związek TGF β1 z postępem uszkodzenia wątroby, równocześnie wskazując na związek progresji pwzw ze zjawiskiem stłuszczenia, insulinooporności i stresu oksydacyjnego wyrażonego wzrostem aktywności surowiczej γ-GTP.
Summary
Transforming growth factor β1 (TGF β1) is main cytokine responsible for progression of fibrosis in chronic liver disease. Factors having impact on progression in chronic viral hepatitis (cvh) are not clearly identified.
The aim of the study was to evaluate factors having influence on progression of fibrosis in cvh.
Hepatic concentration of TGF β1 (Quantikine Immunoassay Kit) was measured in 78 patients: 52 with cvh C, 19 with cvh B, 7 with non alcoholic fatty liver disease and autoimmune hepatitis. The results were analyzed in relation to grade of inflammation and necrotic changes as well as stage of fibrosis in histological analysis of liver bioptates (according to Batt and Ludwig's scale), insulin resistance (HOMA-IR>2.259), serum alanine aminotransferase (ALT), asparagine aminotransferase (AST) and γ-glutamyltranspeptydase (γ-GTP) activity, HCV and HBV viral load, sustained viral response to interferon α therapy in cvh C (SVR), body mass index (BMI), adipose tissue amount and presence of metabolic syndrome (EGIR 1999).
There was correlation between TGF β1 and inflammatory-necrotic process activity on the basis of histological examination (p<0.00001), ALT activity (p=0.0177), degree of fibrosis (p=0.0002). TGF β1 concentration was statistically significantly higher in patients with liver steatosis (p=0.0144), elevated γ-GTP activity (p=0.0296), and insulin resistance (p=0.057).
No statistically significant difference was found in TGF β1 concentration in relation to the rest of analyzed factors.
The results confirmed close correlation between TGF β1 and progression of liver injury as well as correlation between progression of cvh and liver steatosis, insulin resistance and oxydative stress resulting in increase of serum γ-GTP activity.

Wstęp
TGF β1 jest homodimerycznym polipeptydem, szeroko rozpowszechnionym w organizmie, charakteryzującym się dużą aktywnością biologiczną, o wielokierunkowym, zwykle auto- lub parakrynnym mechanizmie działania, obejmującym regulację wzrostu fibroblastów, komórek hematopoetycznych, hepatocytów oraz okołozatokowych komórek Ito (1, 2, 3, 4, 5, 6, 7). W hepatocytach hamuje syntezę DNA, wzrost i proliferację (8, 9, 10). W ostrej i przewlekłej chorobie wątroby jest czynnikiem kontrolującym regenerację komórek wątrobowych (11). Hepatocyty prawidłowej wątroby nie produkują konstytutywnego TGF β1, jego obecność stwierdzana jest w ograniczonej liczbie komórek w marskiej wątrobie (4, 11, 12, 13). W następstwie stanu zapalnego, transformacji nowotworowej lub po częściowej hepatektomii, komórki wątrobowe mogą stać się miejscem produkcji TGF β1 (4, 11, 14, 15). W stanie zapalnym zwiększona ekspresja TGF β1 obserwowana jest w obszarach zapalno-martwiczych (11), miejscach nasilonej odpowiedzi immunologicznej oraz tworzących się przegrodach włóknistych (16).
TGF β1 jest krytycznym czynnikiem w patogenezie rozwoju włóknienia wątroby, silnie indukującym fibrynogenezę w efektorowych komórkach wątrobowego włóknienia – aktywowanych komórkach gwiaździstych, miofibroblastach (12, 16, 17, 18). Komórki gwiaździste są docelowym układem komórkowym dla TGF β1 (6, 7). W następstwie autokrynnej stymulacji komórki te stają się źródłem i celem TGF β1, jednocześnie ulegają transformacji do miofibroblastów (16). TGF β1stymuluje produkcję zewnątrzkomórkowej macierzy, zwiększa w niej ilość kolagenu, glikoprotein, glikozoaminoglikanów (4, 19). W aktywnej chorobie wątroby stwierdzana jest obecność wyznaczników pobudzenia układu TGF β1 w zakresie ligandów, receptorów oraz kaskady sygnalizacyjnej komórkowych przekaźników (19). Ekspresja mRNA i białka TGF β1w tkance wątroby jest współzależna ze stopniem nasilenia włóknienia (11, 12, 19, 20).
W pwzw włóknienie jest procesem określającym szybkość progresji uszkodzenia wątroby. Czynniki wpływające na progresję pwzw nie są jednoznacznie zdefiniowane. W pwzw ważna jest identyfikacja pacjentów z ryzykiem przyśpieszonego włóknienia w celu kwalifikacji ich do leczenia przeciwwirusowego oraz prób terapii hamującej progresję włóknienia (18).
Cel pracy
Celem badania była ocena czynników warunkujących postęp włóknienia w pwzw.
Metoda
Badanie zostało przeprowadzone u 78 chorych diagnozowanych i leczonych w Oddziałach Klinicznych Kliniki Gastroenetreologii i Hepatologii oraz Chorób Zakaźnych Szpitala Uniwersyteckiego w Krakowie. U 52 chorych rozpoznawano pwzw C, 19 pwzw B i 7 niealkoholową chorobę stłuszczeniową wątroby lub autoimmunologiczne zapalenie wątroby. U wszystkich pacjentów wykonano nakłucie gruboigłowe wątroby, ze wskazań wynikających ze standardów postępowania diagnostycznego i terapeutycznego w pzw B i C oraz schorzeniach wątroby o etiologii niezakaźnej. Zabieg nakłucia wątroby wykonywano igłą Hepafix firmy Braun, 1,6 mm. Z uzyskanego bioptatu pobierano fragment o długości 2 mm, który zamrażano i przechowywano w temperaturze -75° C do czasu analizy poziomu TGF β1.
Podstawowa część materiału bioptycznego, została utrwalona w 10% formalinie i kierowana do badania histologicznego w celu określenia nasilenia procesu zapalno-martwiczego, włóknienia miąższu wątroby wg skali Batts'a i Ludwig'a (21) oraz zaawansowania stłuszczenia.
Wycinki do oznaczania stężeń tkankowych TGF β1 były ważone i homogenizowane w buforze fosforanowym o pH 7,1 a następnie wirowanie 4000 obrotów/minutę przez 5 minut. W oddzielonym nadsączu oznaczano stężenie TGF β1 metodą immunoenzymatyczną ELISA z wykorzystaniem zestawu odczynników Quantikine Immunoassay Kit (R&D Systems). W teście zastosowano 96-dołkowe, polistyrenowe płytki, opłaszczone monoklonalnym przeciwciałem, specyficznym dla oznaczanego TGF β1. Do studzienek dodawano standardy oraz nadsącz z homogenatów tkankowych. Po dwugodzinnej inkubacji w temperaturze pokojowej, płytkę kilkakrotnie płukano buforem (Wash Buffer). W kolejnym etapie dodawano specyficznych dla TGF β1 poliklonalnych przeciwciał sprzężonych z peroksydazą chrzanową i ponownie inkubowano przez 2 godziny w temperaturze pokojowej. Po ponownym kilkukrotnym przemyciu, płytki inkubowano przez 30 minut z roztworem tetrametylobenzydyny, związku będącego enzymatycznym substratem peroksydazy chrzanowej. Dodanie roztworu kwasu siarkowego (Stop Solution) zatrzymywało przebieg reakcji barwnej. Wynik odczytywano metodą spektrofotometryczną przy długości fali 450 nm.
W homogenacie bioptatu wątroby oznaczano także stężenie białka metodą Lowry (22). Wyniki przedstawiano w przeliczeniu na masę wycinka tkanki lub stężenie białka. Wartości wyrażono w pg/mg tkanki lub białka.
Stężenie insuliny w surowicy krwi oznaczano metodą immunoradiometryczną zestawem i wg instrukcji producenta BioSource INS-IRMA KIP1251-KIP1254. Insulinooporność rozpoznawano w przypadku wartości HOMA IR powyżej 2,259. Wskaźnik był obliczany wg wzoru:
Oznaczanie aktywności aminotransferazy alaninowej, asparaginowej, γ-glutamylotranspeptydazy, glukozy, cholesterolu całkowitego, lipoprotein o wysokiej i niskiej gęstości, trójglicerydów w surowicy krwi wykonywano zgodnie z procedurą i wykorzystaniem komercyjnych zestawów Cobas firmy Roche.
W oznaczeniu ilościowym HBV, wykorzystywano technologię reakcji PCR czasu rzeczywistego, w oparciu o test COBAS(r) TaqMan(r) HBV, HCV – testu COBAS(r) TaqMan(r) HCV.
W badaniach antropometrycznych określono wagę i wzrost chorego, umożliwiające określenie wskaźnika masy ciała. Przeprowadzono także oznaczenie obwodu tali. Wielkość masy tkanki tłuszczowej określono na podstawie pomiaru fałdu skórno-tłuszczowego okolicy podłopatkowej, nadgrzebieniowej, okolicy mięśnia dwugłowego i trójgłowego ramienia, w oparciu o tablice przeliczeniowe SAEHAN Skinfold Caliper, SH5020, Korea, z uwzględnieniem wieku i płci.
Wyniki oceniano testem analizy wariancji oraz testem χ2 NW. Określono korelację pomiędzy stężeniami TGF β1, w przeliczeniu na masę tkanki bioptatu i stężenie białka miąższu wątroby, w odniesieniu do zaawansowania włóknienia oraz aktywności procesu zapalno-martwiczego, ocenianych nasileniem zmian histopatologicznych oraz aktywnością aminotransferazy alaninowej.
Wyniki
TGF β1 w przeliczeniu na masę bioptatu i stężenia białka miąższu wątroby wykazały współzależność z wartościami wskaźnika korelacji dla procesu zapalno-martwiczego, wynoszącymi odpowiednio r=0,519 i r=0,5187 (p<0,00001, ryc. 1), a także dla fazy włóknienia r=0,41 (p=0,0002) oraz r=0,43 (p=0,00009, ryc. 2). W przypadku oceny aktywności aminotransferazy alaninowej korelację wykazano tylko w oznaczeniu TGF β1 w przeliczeniu na masę tkanki próbki r=0,2787 (p=0,0177).
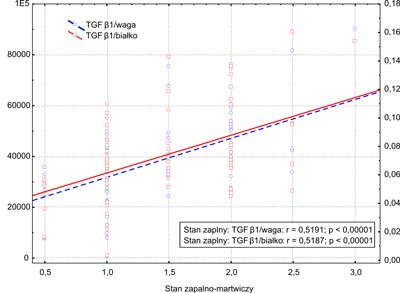
Ryc. 1. Korelacja aktywności stanu zapalno-martwiczego miąższu wątroby oraz TGF β1 oznaczanego w pikogramach na miligram próbki (linia przerywana: TGF β1/waga) lub w pikogramach na miligram białka miąższu wątroby (linia ciągła: TGF β1/białko).
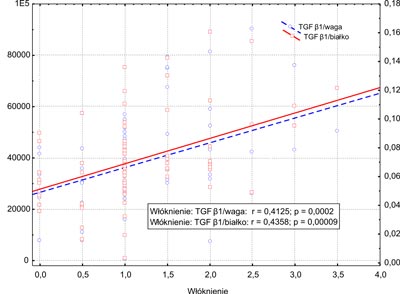
Ryc. 2. Korelacja stopnia włóknienia miąższu wątroby oraz TGF β1 oznaczanego w pikogramach na miligram próbki, linia przerywana: TGF β1/waga) lub w pikogramach na miligram białka miąższu wątroby (linia ciągła: TGF β1/białko).
Podobnie jak dla całej grupy badanych chorych w przypadku pzw C i B w teście analizy wariancji stwierdzono zróżnicowanie zależne od stanu zapalno-martwiczego z wartościami prawdopodobieństwa testowego, wynoszącymi p= 0,0072 oraz p= 0,0129, fazy włóknienia tylko w przypadku pzw C, p= 0,0002. Statystycznie wyższe wartości TGF β1 obserwowane były wśród chorych z pzw C z obecnością stłuszczenia miąższu wątroby (p=0,0144), zwiększoną aktywnością γ-glutamylotranspeptydazy (γ-GTP) (p=0,0296) oraz na granicy istotności statystycznej z insulinoopornością (p=0,057, ryc. 3).
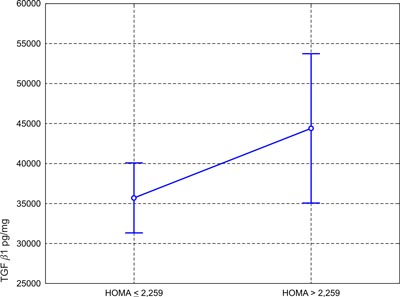
Ryc. 3. Zróżnicowanie stężenia TGF β1, w teście analizy wariancji u chorych z pzw C, B i grupie kontrolnej w zależności od braku lub obecności insulinooporności. Test analizy wariancji, p= 0,0574.
Powyżej zamieściliśmy fragment artykułu, do którego możesz uzyskać pełny dostęp.
Mam kod dostępu
- Aby uzyskać płatny dostęp do pełnej treści powyższego artykułu albo wszystkich artykułów (w zależności od wybranej opcji), należy wprowadzić kod.
- Wprowadzając kod, akceptują Państwo treść Regulaminu oraz potwierdzają zapoznanie się z nim.
- Aby kupić kod proszę skorzystać z jednej z poniższych opcji.
Opcja #1
29 zł
Wybieram
- dostęp do tego artykułu
- dostęp na 7 dni
uzyskany kod musi być wprowadzony na stronie artykułu, do którego został wykupiony
Opcja #2
69 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 30 dni
- najpopularniejsza opcja
Opcja #3
129 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 90 dni
- oszczędzasz 78 zł
Piśmiennictwo
1. Cross D, Cambier JC: Transforming growth factor beta 1 has differential effects on B cell proliferation and activation antigen expression. J Immunol 1990; 144(2): 432-9.
2. Ignotz RA, Massagué J: Transforming growth factor-beta stimulates the expression of fibronectin and collagen and their incorporation into the extracellular matrix. J Biol Chem 1986; 261(9): 4337-45.
3. Oberhammer FA, Pavelka M, Sharma S et al.: Induction of apoptosis in cultured hepatocytes and in regressing liver by transforming growth factor beta 1. Proc Natl Acad Sci USA 1992; 89(12): 5408-12.
4. Bedossa P, Peltier E, Terris B et al.: Transforming growth factor-beta 1 (TGF-beta 1) and TGF-beta 1 receptors in normal, cirrhotic, and neoplastic human livers. Hepatology 1995; 21(3): 760-6.
5. Kelin A: Molekularne podstawy regulacji hormonalnej (Sygnalizacja międzykomórkowa i wewnątrzkomórkowa). Copyright by Wydział Biotechnologii UJ Kraków 2002.
6. Bachem MG, Meyer D, Melchior R et al.: Activation of rat liver perisinusoidal lipocytes by transforming growth factors derived from myofibroblastlike cells. A potential mechanism of self perpetuation in liver fibrogenesis. J Clin Invest 1992; 89(1): 19-27.
7. Friedman SL, Arthur MJ: Activation of cultured rat hepatic lipocytes by Kupffer cell conditioned medium. Direct enhancement of matrix synthesis and stimulation of cell proliferation via induction of platelet-derived growth factor receptors. J Clin Invest 1989; 84(6): 1780-5.
8. Carr BI, Hayashi I, Branum EL et al.: Inhibition of DNA synthesis in rat hepatocytes by platelet-derived type beta transforming growth factor. Cancer Res 1986; 46(5): 2330-4.
9. Jakowlew SB, Mead JE, Danielpour D et al.: Transforming growth factor-beta (TGF-beta) isoforms in rat liver regeneration: messenger RNA expression and activation of latent TGF-beta. Cell Regul 1991; 2(7): 535-48.
10. Russell WE, Coffey RJ Jr, Ouellette AJ et al.: Type beta transforming growth factor reversibly inhibits the early proliferative response to partial hepatectomy in the rat. Proc Natl Acad Sci USA 1988; 85(14): 5126-30.
11. Milani S, Herbst H, Schuppan D et al.: Transforming growth factors beta 1 and beta 2 are differentially expressed in fibrotic liver disease. Am J Pathol 1991; 139(6): 1221-9.
12. Castilla A, Prieto J, Fausto N: Transforming growth factors beta 1 and alpha in chronic liver disease. Effects of interferon alfa therapy. N Engl J Med 1991; 324(14): 933-40.
13. Roulot D, Durand H, Coste T et al.: Quantitative analysis of transforming growth factor beta 1 messenger RNA in the liver of patients with chronic hepatitis C: absence of correlation between high levels and severity of disease. Hepatology 1995; 21(2): 298-304.
14. Rizzino A: Understanding the roles of growth factors in carcinogenesis: modulation of autocrine growth control by differentiation. Int J Dev Biol 1993; 37(1): 61-5.
15. Schirmacher P, Rogler CE, Dienes HP: Current pathogenetic and molecular concepts in viral liver carcinogenesis. Virchows Arch B Cell Pathol Incl Mol Pathol 1993; 63(2): 71-89.
16. Bayer EM, Herr W, Kanzler S et al.: Transforming growth factor-beta1 in autoimmune hepatitis: correlation of liver tissue expression and serum levels with disease activity. J Hepatol 1998; 28(5): 803-11.
17. Suzuki S, Tanaka Y, Orito E et al.: Transforming growth factor-beta-1 genetic polymorphism in Japanese patients with chronic hepatitis C virus infection. J Gastroenterol Hepatol 2003; 18(10): 1139-43.
18. Schuppan D, Krebs A, Bauer M et al.: Hepatitis C and liver fibrosis. Cell Death Differ 2003; 10 Suppl 1: S59-67.
19. Calabrese F, Valente M, Giacometti C et al.: Parenchymal transforming growth factor beta-1: its type II receptor and Smad signaling pathway correlate with inflammation and fibrosis in chronic liver disease of viral etiology. J Gastroenterol Hepatol 2003; 18(11): 1302-8.
20. Annoni G, Weiner FR, Zern MA: Increased transforming growth factor-beta 1 gene expression in human liver disease. J Hepatol 1992; 14(2-3): 259-64.
21. Batts KP, Ludwig J: Chronic hepatitis, an update on terminology and reporting. Am J Surg Pathol 1995; 19(12): 1409-1417.
22. Lowry OH, Rosebrough NJ, Farr AL et al.: Protein measurement with the Folin phenol reagent. J Biol Chem 1951; 193(1): 265-75.
23 Anscher MS, Crocker IR, Jirtle RL: Transforming growth factor-beta 1 expression in irradiated liver. Radiat Res 1990; 12, 2(1): 77-85.
24. Nelson DR, Gonzalez-Peralta RP, Qian K et al.: Transforming growth factor-beta 1 in chronic hepatitis C. J Viral Hepat 1997; 4(1): 29-35.
25. Kinnman N, Andersson U, Hultcrantz R: In situ expression of transforming growth factor-beta1-3, latent transforming growth factor-beta binding protein and tumor necrosis factor-alpha in liver tissue from patients with chronic hepatitis C. Scand J Gastroenterol 2000; 35(12): 1294-300.
26. Czaja MJ, Flanders KC, Biempica L et al.: Expression of tumor necrosis factor-alpha and transforming growth factor-beta 1 in acute liver injury. Growth Factors 1989; 1(3): 219-26.
27. Miwa Y, Harrison PM, Farzaneh F et al.: Plasma levels and hepatic mRNA expression of transforming growth factor-beta1 in patients with fulminant hepatic failure. J Hepatol 1997; 27(5): 780-8.
28. Blonsky JJ, Harrison SA: Review article: nonalcoholic fatty liver disease and hepatitis C virus – partners in crime. Aliment Pharmacol Ther 2008; 27(10): 855-65.
29. Gabriel A, Ziólkowski A, Radlowski P et al.: Hepatocyte steatosis in HCV patients promotes fibrosis by enhancing TGF-beta liver expression. Hepatol Res 2008; 38(2): 141-6.
30. Kakumu S, Ito Y, Wakita T et al.: Effects of transforming growth factor-beta 1 against the inhibitory action of interferon on DNA synthesis and viral replication in hepatitis B virus DNA-transfected cell. J Med Virol 1992; 38(1): 62-6.
31. Muzzi A, Leandro G, Rubbia-Brandt L et al.: Swiss Hepatitis C Cohort Study. Insulin resistance is associated with liver fibrosis in non-diabetic chronic hepatitis C patients. J Hepatol 2005; 42(1): 41-6.
32. Svegliati-Baroni G, Ridolfi F, Di Sario A et al.: Insulin and insulin-like growth factor-1 stimulate proliferation and type I collagen accumulation by human hepatic stellate cells: differential effects on signal transduction pathways. Hepatology 1999; 29(6): 1743-51.
33. Lee DH, Blomhoff R, Jacobs DR Jr: Is serum gamma glutamyltransferase a marker of oxidative stress? Free Radic Res 2004; 38(6): 535-9.
34. Leandro G, Mangia A, Hui J et al.: HCV Meta-Analysis (on) Individual Patients' Data Study Group. Relationship between steatosis, inflammation, and fibrosis in chronic hepatitis C: a meta-analysis of individual patient data. Gastroenterology 2006;130(6): 1636-42.
35. Ratziu V, Trabut JB, Poynard T: Fat, diabetes, and liver injury in chronic hepatitis C. Curr Gastroenterol Rep 2004; 6(1): 22-9.
