Marta Kula, *Mirosława Krauze-Baranowska, Natalia Adamczuk
Urolityny – aktywne biologicznie produkty metabolizmu elagotanin
Urolithins – biologically active products of ellagitannin metabolism
Katedra i Zakład Farmakognozji, Wydział Farmaceutyczny, Gdański Uniwersytet Medyczny
Kierownik Katedry: prof. dr hab. n. farm. Mirosława Krauze-Baranowska
Streszczenie
Elagotaniny są grupą metabolitów wtórnych obecnych w orzechach i wielu owocach, takich jak: granaty, maliny, truskawki, jeżyny i żurawiny. Aktywnymi biologicznie produktami metabolizmu elagotanin w organizmie człowieka, w zależności od kompozycji mikrobiomu jelitowego, są urolityny A, B, C lub D. W pracy scharakteryzowano dotychczasowe wyniki prac nad aktywnością biologiczną urolityn, m.in.: przeciwzapalną, antyoksydacyjną, chemoprewencyjną, modulacją receptora estrogenowego i neuroprotekcyjną. Wskazano na pozytywne efekty urolityn m.in. w leczeniu nieswoistego zapalenia jelit, a także chorób układu sercowo-naczyniowego. Ponadto opisano korzystne właściwości elagotanin w zespole metabolicznym, w tym działanie przeciwglikemiczne i neuroprotekcyjne.
Summary
Ellagitannins are a group of secondary metabolites found in nuts and many fruits such as pomegranates, raspberries, strawberries, blackberries and cranberries. The biologically active products of ellagitannin metabolism in the human body, depending on the composition of the intestinal microbiome, are urolithins A, B, C or D. The work presents the results of research on the biological activity of urolithins, including anti-inflammatory, antioxidant, chemopreventive, modulation of estrogenic receptors and neuroprotective activity. The positive effect of urolithins in inflammatory bowel diseases as well as in diseases of the cardiovascular system is characterized. In addition, the beneficial properties of urolithins in metabolic syndrome, including an anti-glycemic and neuroprotective effect, have been described.

Urolityny – aktywne biologicznie produkty metabolizmu elagotanin
Elagotaniny są estrami kwasu heksahydroksydifenylowego (HHDP) oraz monocukrów (glukoza) (1-5). Jedną z właściwości elagotanin jest ich hydroliza w środowisku kwaśnym i zasadowym z uwolnieniem kwasu heksahydroksydifenylowego, który przekształca się (laktonizacja) w kwas elagowy (1, 5, 6). Elagotaniny z ugrupowaniem HHDP, posiadającym wiązanie C-C między fragmentami galoilowymi, estryfikującymi szkielet glukozy, są szeroko rozpowszechnione w świecie roślinnym. Podstawowe biarylowe wiązanie C-C obejmuje przede wszystkim galoilowe grupy estryfikujące cząsteczkę glukozy w pozycjach węgli C -2,3- i -4,6-; spotykane są również wiązania w pozycjach C -1,6-, -1,3- lub -3,6- i/lub -2,4- (1, 7). Do grupy elagotanin zaliczane są związki o charakterze monomerów (np. nufaryna A), oligomerów (nufaryna E) oraz polimerów (10).
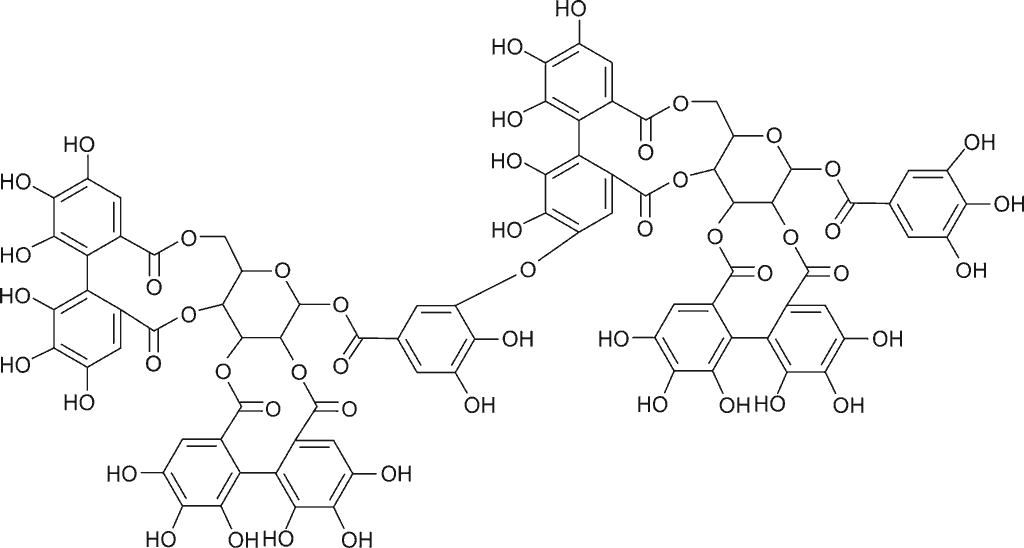
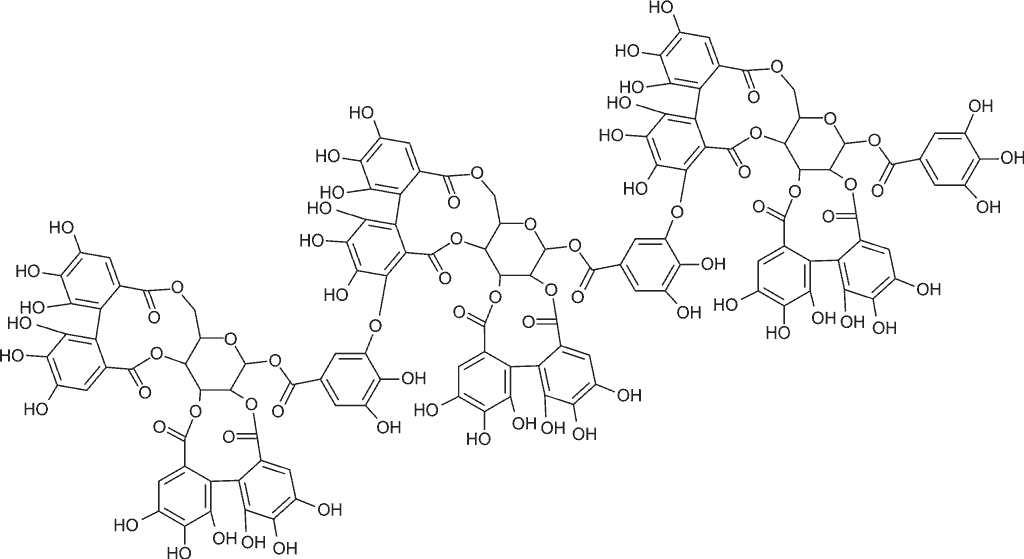
Elagotaniny o charakterze oligomerów dominują w owocach różnych odmian uprawowych maliny właściwej Rubus idaeus (8); są to sangwina H-6 (ryc. 1) oraz lambertianina C (ryc. 2) (3). W świecie roślinnym obok owoców maliny bogatym źródłem elagotanin są również inne owoce gatunków z rodziny Rosaceae: owoce truskawki, jeżyny oraz róży (1, 9); z rodziny Saxifragaceae: owoce czarnej porzeczki; z rodziny Ericaceae: owoce czarnej jagody i żurawiny. Wyjątkowo bogate w elagotaniny są owoce granatu i winorośli. Ponadto elagotaniny w wysokich stężeniach są obecne w owocach i nasionach o zdrewniałej owocni – orzechach włoskich, pistacjach, orzechach pekan (nasiona orzesznika jadalnego), orzechach nerkowca, a także w orzechach dębu i nasionach kasztanowca (10). Elagotaniny mogą być wytrawiane z drewna np. dębowego, z którego wykonane są beczki, i przechodzić do dojrzewającego w nich wyrobu alkoholowego (wino).
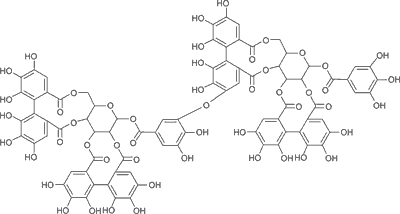
Ryc. 1. Wzór strukturalny sangwiny H-6
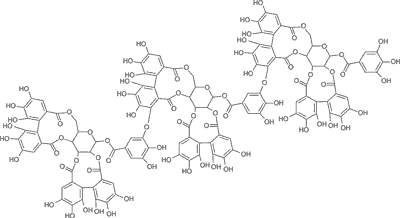
Ryc. 2. Wzór strukturalny lambertaniny C
Elagotaniny wykazują prozdrowotne właściwości i znaczącą aktywność biologiczną: przeciwzapalną, przeciwmiażdżycową, przeciwzakrzepową i antyangiogenną, stąd klasyfikowane są jako nutraceutyki. Badania potwierdziły związek między spożywaniem żywności bogatej w elagotaniny (orzechy włoskie, owoce granatu) a poprawą funkcji układu sercowo-naczyniowego. Obecnie na rynku dostępnych jest wiele produktów z wyciągami roślinnymi, zawierającymi elagotaniny i kwas elagowy (10).
Ze względu na szerokie spektrum aktywności biologicznej elagotanin i kwasu elagowego (11) w licznych pracach badano ich absorpcję z przewodu pokarmowego oraz dystrybucję w organizmie (12-28).
Metabolizm elagotanin oraz ich biodostępność
W badaniach na modelach zwierzęcych oraz w badaniach u ludzi wykazano, że absorpcja elagotanin z przewodu pokarmowego jest ograniczona (12-15). U szczurów długotrwale spożywających znaczne ilości elagotanin jedynie 3-6% było obecnych w formie niezmetabolizowanej w moczu i osoczu (12), podczas gdy u ludzi nie stwierdzono ich obecności w żadnym z płynów ustrojowych (13-15). Ustalono natomiast, że większość elagotanin jest metabolizowana przez mikroflorę jelitową do związków zwanych urolitynami.
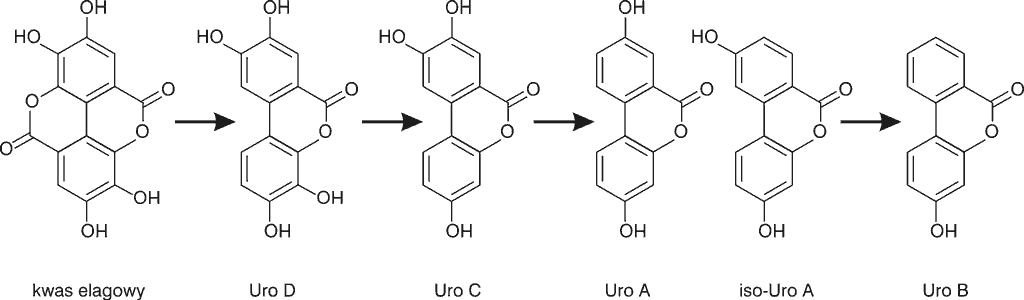
Po spożyciu, elagotaniny (ET) ulegają degradacji w kwaśnym środowisku przewodu pokarmowego, hydrolizując z uwolnieniem kwasu heksahydroksydifenowego (HHDP), który na drodze laktonizacji przekształcany jest w kwas elagowy (EA) (16). Badania płynnej treści jelitowej od pacjentów z ileostomią sugerują, że ok. 80% sangwiny H-6 ulega nahydrolizie do kwasu elagowego w żołądku oraz w jelicie cienkim (15). Ponadto 80% wolnego kwasu elagowego jest przekształcane przez mikroflorę jelita grubego w urolityny, poprzez otwarcie i dekarboksylację jednego z pierścieni laktonowych i degradację kolejnych grup hydroksylowych. W wyniku tego procesu powstają urolityny tetrahydroksylowe (urolityna D, Uro D), trihydroksylowe (urolityna C, Uro C), dihydroksylowe (urolityna A, Uro A i izourolityna A iso-Uro A) oraz monohydroksylowe (urolityna B, Uro B), które są pochodnymi 6H-dibenzo-β-D-piran-6-onu (ryc. 3) (13, 15-21).
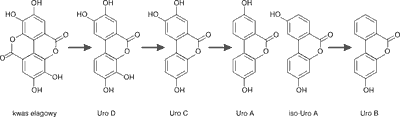
Ryc. 3. Schemat konwersji kwasu elagowego do urolityn (17)
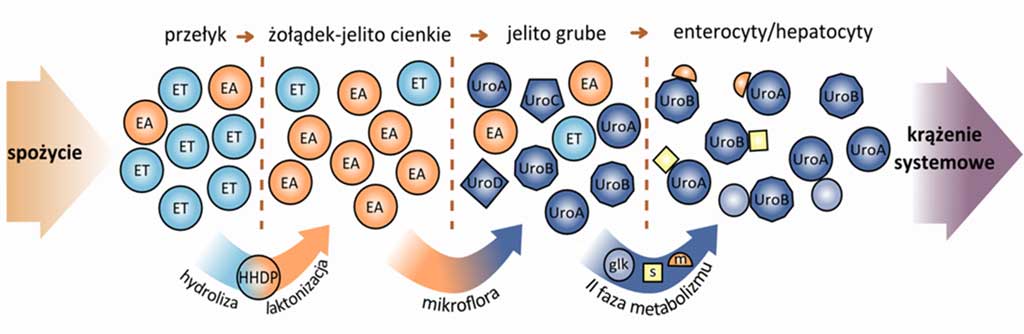
Po absorpcji w jelicie grubym w II fazie metabolizmu urolityny ulegają metylacji, glukuronidacji i sulfonacji, które zachodzą w ścianie jelita, bądź w wątrobie (ryc. 4) (15, 17, 18). Obecnie uważa się, że urolityny są formą biodostępną elagotanin, ulegającą absorpcji i dystrybucji do tkanek in vivo (15, 17, 19-21). W oparciu o badania na modelach zwierzęcych u myszy i świni iberyjskiej stwierdzono obecność urolityn w hepatocytach (19, 20), woreczku żółciowym oraz żółci wydzielanej do dwunastnicy (19), co sugeruje, że urolityny i ich glukuronidy są włączane do obiegu jelitowo-wątrobowego.
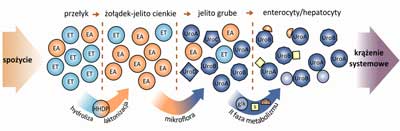
Ryc. 4. Metabolizm elagotanin i kwasu elagowego
Obecność urolityn wykazano również w innych tkankach (20, 21). Urolityny (Uro A i jej glukuronid) wykryto w niskich stężeniach w gruczole krokowym i nerkach u myszy (20) oraz w gruczole krokowym u ludzi. Wyniki biopsji prostaty wykazały obecność glukuronidu urolityny A w stężeniu 2 ng/g i śladowe ilości glukuronidu urolityny B (21). Inne tkanki badane w zakresie akumulacji urolityn i ich pochodnych w modelach zwierzęcych obejmowały tkankę mięśniową, tłuszczową i sercową, niemniej jednak w żadnej z nich nie ujawniono ich obecności (17, 19).
Urolityny przechodzą do krwi i są wydalane z moczem. W płynach ustrojowych u ludzi wykryto obecność Uro A-D oraz szeregu ich glukuronidowych, sulfonowych i metylowych pochodnych, z dominującym glukuronidem urolityny A (ryc. 4) (12-15, 17, 19, 22-24). Metabolity elagotanin są wykrywane w moczu po 1-2 dniach od spożycia produktów będących ich źródłem, takich jak: maliny, truskawki, orzechy i owoce granatu. Zarówno w moczu, jak i we krwi obserwowane jest duże zróżnicowanie osobnicze w zakresie oznaczonych stężeń urolityn (14, 15, 19, 25). Uważa się, że skład flory bakteryjnej jelita grubego wpływa na różnice w stężeniach i produkcji poszczególnych urolityn (14, 15, 25, 26).
Na podstawie dotychczasowych badań u ludzi można wyróżnić trzy grupy różniące się metabolizmem elagotanin: grupę produkującą głównie urolitynę A (25-80%), grupę metabolizującą głównie do urolityny B (10-50%) oraz grupę, u której nie obserwuje się produkcji urolityn (5-25%). Przynależność do danej grupy nie jest skorelowana z płcią, wiekiem, BMI, ilością oraz źródłem spożytych elagotanin. Obserwowano natomiast, że Uro B częściej produkują pacjenci z przewlekłymi schorzeniami, takimi jak: zespół metaboliczny, nowotwór jelita grubego czy dysbioza jelitowa (27). U 10-50% ludzi, oprócz urolityny A, obserwowano produkcję jej izomeru – izourolityny A (iso-Uro A) (28).
Aktywność biologiczna urolityn
W świetle ostatnich wyników badań przyjmuje się, że to urolityny, a nie formy natywne elagotanin, są w głównej mierze odpowiedzialne za obserwowane efekty biologiczne in vivo. Skutkuje to wzrostem liczby prac dotyczących ich aktywności biologicznej (14, 28-60).
Aktywność antyoksydacyjna
Dotychczasowe wyniki badań aktywności antyoksydacyjnej urolityn są rozbieżne. W testach z użyciem DPPH i redukcji rodnika ABTS [2,2’-azobis(3-etylobenzotiazolino-6-sulfonian)] wykazano, że w porównaniu z elagotaninami urolityna A charakteryzuje się bardzo niską aktywnością antyoksydacyjną, ok. 42 razy słabszą w teście DPPH oraz aż 3500 razy słabszą w teście ABTS (14). Podobnie w badaniach Larrosa i wsp. (29) wartości IC50 urolityn w szeregu testów oksydacyjnych (m.in. z DPPH i NADH) wynosiły powyżej 100 μM, jakkolwiek urolityny, a w szczególności Uro A w teście ORAC (ang. oxygen radical absorbance capacity), badającym zdolność pochłaniania reaktywnych form tlenu, wykazywały aktywność podobną do kwasu askorbinowego. Wyniki te są zgodne z innymi badaniami porównującymi właściwości antyutleniające związków fenolowych (m.in. flawonoli, flawan-3-oli, kwasów fenolowych) i ich metabolitów metodą ORAC, spośród których urolityna A była jednym z najsilniejszych antyoksydantów (30).
Przeciwutleniający potencjał urolityn badano również w komórkowym teście antyoksydacyjnym. W badaniach na linii HL-60 stwierdzono, że urolityny C i D są najsilniejszymi antyoksydantami z wartościami IC50 wynoszącymi odpowiednio 0,16 i 0,33 μM, silniejszymi niż kwas elagowy (IC50 1,1 μM) i witamina C (IC50 1,9 μM). Urolityna A, pomimo że charakteryzowała się niższym potencjałem antyoksydacyjnym (IC50 13,6 μM), to odpowiadała zakresowi stężeń osiąganymi in vivo, podczas gdy urolityna B nie wykazywała aktywności antyoksydacyjnej. Zaletą testów antyoksydacyjnych przeprowadzonych na komórkach, w przeciwieństwie do testów enzymatycznych, jest uwzględnienie transportu przez błony i możliwość oceny biodostępności badanych związków (31).
Wyniki przeprowadzonych eksperymentów sugerują, że urolityny charakteryzują się wyższym potencjałem antyoksydacyjnym niż początkowo sądzono, przy czym obserwuje się dużą zmienność wyników w zależności od zastosowanej metody badawczej, co wymaga dalszych badań w warunkach in vivo (32).
Aktywność przeciwzapalna
Powyżej zamieściliśmy fragment artykułu, do którego możesz uzyskać pełny dostęp.
Mam kod dostępu
- Aby uzyskać płatny dostęp do pełnej treści powyższego artykułu albo wszystkich artykułów (w zależności od wybranej opcji), należy wprowadzić kod.
- Wprowadzając kod, akceptują Państwo treść Regulaminu oraz potwierdzają zapoznanie się z nim.
- Aby kupić kod proszę skorzystać z jednej z poniższych opcji.
Opcja #1
29 zł
Wybieram
- dostęp do tego artykułu
- dostęp na 7 dni
uzyskany kod musi być wprowadzony na stronie artykułu, do którego został wykupiony
Opcja #2
69 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 30 dni
- najpopularniejsza opcja
Opcja #3
129 zł
Wybieram
- dostęp do tego i pozostałych ponad 7000 artykułów
- dostęp na 90 dni
- oszczędzasz 78 zł
Piśmiennictwo
1. Cerdá B, Periago P, Espín JC i wsp. Identification of urolithin A as a metabolite produced by human colon microflora from ellagic acid and related compounds. J Agric Food Chem 2005; 53:5571-6.
2. Puupponen-Pimiä R, Seppänen-Laakso T, Kankainen M i wsp. Effects of ellagitannin-rich berries on blood lipids, gut microbiota, and urolithin production in human subjects with symptoms of metabolic syndrome. Mol Nutr Food Res 2013; 57:2258-63.
3. Cho H, Jung H, Lee H i wsp. Chemopreventive activity of ellagitannins and their derivatives from black raspberry seeds on HT-29 colon cancer cells. Food Funct 2015; 6:1675-83.
4. González-Sarrías A, Espín JC, Tomás-Barberán FA i wsp. Gene expression, cell cycle arrest and MAPK signalling regulation in Caco-2 cells exposed to ellagic acid and its metabolites, urolithins. Mol Nutr Food Res 2009; 53:686-98.
5. Wyzgoski FJ, Liladhar P, Rinaldi PL i wsp. Modeling relationships among active components in black raspberry (Rubus occidentalis L.) fruit extracts using high-resolution1H nuclear magnetic resonance (NMR) spectroscopy and multivariate statistical analysis. J Agric Food Chem 2010; 58:3407-14.
6. Kim LS, Youn SH, Kim JY. Comparative study on antioxidant effects of extracts from Rubus coreanus and Rubus occidentalis. J Korean Soc Food Sci Nutr 2014; 43:1357-62.
7. Ożarowski A, Jaroniewski W. Rośliny lecznicze i ich praktyczne zastosowanie. Instytut Wydawniczy Związków Zawodowych, Warszawa 1987.
8. Kalt W, Forney CF, Martin A i wsp. Antioxidant capacity, vitamin C, and anthocyanins after storage of small fruits. J Agric Food Chem 1999; 47:4638-44.
9. Bialonska D, Ramnani P, Kasimsetty SG i wsp. The influence of pomegranate by-product and punicalagins on selected groups of human intestinal microbiota. Int J Food Microbiol 2010; 140:175-82.
10. Lipińska L, Klewicka E, Sójka M. Structure, occurrence and biological activity of ellagitannins: a general review. Acta Sci Pol 2014; 13:289-99.
11. Majdan M. Badania składu chemicznego Rubus idaeus i Rubus occidentalis z uwzględnieniem aktywności farmakologicznej. Katedra i Zakład Farmakognozji, Gdański Uniwersytet Medyczny, Gdańsk 2013.
12. Cerdá B, Llorach R, Cerón JJ i wsp. Evaluation of the bioavailability and metabolism in the rat of punicalagin, an antioxidant polyphenol from pomegranate juice. Eur J Nutr 2003; 42:18-28.
13. Cerdá B, Tomás-Barberán FA, Espín JC. Metabolism of antioxidant and chemopreventive ellagitannins from strawberries, raspberries, walnuts, and oak-aged wine in humans: Identification of biomarkers and individual variability. J Agric Food Chem 2005; 53:227-35.
14. Cerdá B, Espín JC, Perra S i wsp. The potent in vitro antioxidant ellagitannins from pomegranate juice are metabolised into bioavailable but poor antioxidant hydroxy-6H-dibenzopyran-6-one derivatives by the colonic microflora of healthy humans. Eur J Nutr 2004; 43:205-20.
15. González-Barrio R, Edwards CA, Crozier A. Colonic catabolism of ellagitannins, ellagic acid, and raspberry anthocyanins: in vivo and in vitro studies. Drug Metab Dispos 2011; 39:1680-8.
16. Landete JM. Ellagitannins, ellagic acid and their derived metabolites: A review about source, metabolism, functions and health. Food Res Int 2011; 44:1150-60.
17. Mena P, Bruni R, Calani L i wsp. Chapter 6 – Bioactivation of high-molecular-weight polyphenols by the gut microbiome, in diet-microbe interactions in the gut. Academic Press, San Diego 2015; 73-101.
18. Zhou BH, Jin L, Tu J i wsp. Metabolites of tannins in Punica granatum L. husk in rat urine. J Chinese Pharm Sci 2014; 49:821-4.
19. Espín JC, González-Barrio R, Cerdá B. Iberian pig as a model to clarify obscure points in the bioavailability and metabolism of ellagitannins in humans. J Agric Food Chem 2007; 55:10476-85.
20. Seeram NP, Aronson WJ, Zhang Y i wsp. Pomegranate ellagitannin-derived metabolites inhibit prostate cancer growth and localize to the mouse prostate gland. J Agric Food Chem 2007; 55:7732-7.
21. González-Sarrías A, Gimènez-Bastida JA, García-Conesa MT. Occurrence of urolithins, gut microbiota ellagic acid metabolites and proliferation markers expression response in the human prostate gland upon consumption of walnuts and pomegranate juice. Mol Nutr Food Res 2010; 54:311-22.
22. Tulipani S, Urpi-Sarda M, García-Villalba R i wsp. Urolithins are the main urinary microbial-derived phenolic metabolites discriminating a moderate consumption of nuts in free-living subjects with diagnosed metabolic syndrome. J Agric Food Chem 2012; 60:8930-40.
23. Truchado P, Larossa M, García-Conesa MT i wsp. Strawberry processing does not affect the production and urinary excretion of acid metabolites, in humans. J Agric Food Chem 2012; 60:5749-54.
24. Mertens-Talcott SU, Jilma-Stohlawetz P, Rios J i wsp. Absorption, metabolism, and antioxidant effects of pomegranate (Punica granatum L.) polyphenols after ingestion of a standardized extract in healthy human volunteers. J Agric Food Chem 2006; 54:8956-61.
25. García-Muñoz C, Hernández L, Pèrez A i wsp. Diversity of urinary excretion patterns of main ellagitannins’ colonic metabolites after ingestion of tropical highland blackberry (Rubus adenotrichus) juice. Food Res Int 2014; 55:161-9.
26. Selma MV, Espín JC, Tomás-Barberán FA. Interaction between phenolics and gut microbiota: Role in human health. J Agric Food Chem 2009; 57:6485-501.
27. Tomás-Barberán FA, García-Villalba R, González-Sarrías A i wsp. Ellagic acid metabolism by human gut microbiota: Consistent observation of three urolithin phenotypes in intervention trials, independent of food source, age, and health status. J Agric Food Chem 2014; 62:6535-8.
28. González-Sarrías A, Núñez-Sánchez MA, García-Villalba R i wsp. Antiproliferative activity of the ellagic acid-derived gut microbiota isourolithin A and comparison with its urolithin A isomer: the role of cell metabolism. Eur J Nutr 2015; 1-11.
29. Larrosa M, González-Sarrías A, Yáñez-Gascón MJ i wsp. Anti-inflammatory properties of a pomegranate extract and its metabolite urolithin-A in a colitis rat model and the effect of colon inflammation on phenolic metabolism. J Nutr Biochem 2010; 21:717-25.
30. Ishimoto H, Tai A, Yoshimura M. Antioxidative properties of functional polyphenols and their metabolites assessed by an ORAC assay. Biosci Biotechnol Biochem 2012; 76:395-9.
31. Dobroslawa B, Kasimsetty SG, Khan SI i wsp. Urolithins, intestinal microbial metabolites of pomegranate ellagitannins, exhibit potent antioxidant activity in a cell-based assay. J Agric Food Chem 2009; 57:10181-6.
32. Kallio T, Kallio J, Jaakkola M i wsp. Urolithins display both antioxidant and pro-oxidant activities depending on assay system and conditions. J Agric Food Chem 2013; 61:10720-9.
33. Gimènez-Bastida JA, Larossa M, González-Sarrías A i wsp. Intestinal ellagitannin metabolites ameliorate cytokine-induced inflammation and associated molecular markers in human colon fibroblasts. J Agric Food Chem 2012; 60:8866-76.
34. González-Sarrías A, Larossa M, Tomás-Barberán FA i wsp. NF-κB-dependent anti-inflammatory activity of urolithins, gut microbiota ellagic acid-derived metabolites, in human colonic fibroblasts. Br J Nutr 2010; 104:503-12.
35. Piwowarski JP, Kiss AK, Granica S i wsp. Urolithins, gut microbiota-derived metabolites of ellagitannins, inhibit LPS-induced inflammation in RAW 264.7 murine macrophages. Mol Nutr Food Res 2015; 59(11):2168-77.
36. Piwowarski JP, Granica S, Zwierzyńska M i wsp. Role of human gut microbiota metabolism in the anti-inflammatory effect of traditionally used ellagitannin-rich plant materials. J Ethnopharmacol 2014; 155:801-9.
37. Piwowarski JP, Granica S, Kiss AK. Influence of gut microbiota-derived ellagitannins metabolites urolithins on pro-inflammatory activities of human neutrophils. Planta Med 2014; 80:887-95.
38. Kiss AK, Granica S, Stolarczyk M i wsp. Epigenetic modulation of mechanisms involved in inflammation: Influence of selected polyphenolic substances on histone acetylation state. Food Chem 2012; 131:1015-20.
39. Vicinanza R, Zhang Y, Henning SM i wsp. Pomegranate juice metabolites, ellagic acid and urolithin a, synergistically inhibit androgen-independent prostate cancer cell growth via distinct effects on cell cycle control and apoptosis. Evid Based Complem Altern Med 2013.
40. Kasimsetty SG, Bialonska D, Reddy MK i wsp. Effects of pomegranate chemical constituents/intestinal microbial metabolites on CYP1B1 in 22Rv1 prostate cancer cells. J Agric Food Chem 2009; 57:10636-44.
41. Sánchez-González C, Ciudad CJ, Noè V i wsp. Walnut polyphenol metabolites, urolithins A and B, inhibit the expression of the prostate-specific antigen and the androgen receptor in prostate cancer cells. Food and Funct 2014; 5:2922-30.
42. Giorgio C, Mena P, Del Rio D i wsp. The ellagitannin colonic metabolite urolithin D selectively inhibits EphA2 phosphorylation in prostate cancer cells. Mol Nutr Food Res 2015; 59(11):2155-67.
43. Sánchez-González C, Ciudad CJ, Izquierdo-Pulido M i wsp. Urolithin A causes p21 up-regulation in prostate cancer cells. Eur J Nutr, 2016; 55:1099-112.
44. Nuñez-Sánchez MA, García-Villalba R, Monedero-Saiz T i wsp. Targeted metabolic profiling of pomegranate polyphenols and urolithins in plasma, urine and colon tissues from colorectal cancer patients. Mol Nutr Food Res 2014; 58:1199-211.
45. González-Sarrías A, Gimènez-Bastida JA, Nuñez-Sán-chez MA i wsp. Phase-II metabolism limits the antiproliferative activity of urolithins in human colon cancer cells. Eur J Nutr 2014; 53:853-64.
46. Kasimsetty SG, Bialonska D, Reddy MK i wsp. Colon cancer chemopreventive activities of pomegranate ellagitannins and Urolithins. J Agric Food Chem 2010; 58(4):2180-7.
47. Sharma M, Li L, Celver J i wsp. Effects of fruit ellagitannin extracts, ellagic acid, and their colonic metabolite, urolithin A, on Wnt signaling. J Agric Food Chem 2010; 58:3965-9.
48. González-Sarrías A, Tomè-Carneiro J, Ballesia A i wsp. The ellagic acid-derived gut microbiota metabolite, urolithin A, potentiates the anticancer effects of 5-fluorouracil chemotherapy on human colon cancer cells. Food and Funct 2015; 6:1460-9.
49. Qiu Z, Zhou B, Jin L i wsp. In vitro antioxidant and antiproliferative effects of ellagic acid and its colonic metabolite, urolithins, on human bladder cancer T24 cells. Food Chem Toxicol 2013; 59:428-37.
50. Adams LS, Zhang Y, Seeram NP i wsp. Pomegranate ellagitannin-derived compounds exhibit antiproferative and antiaromatase activity in breast cancer cells in vitro. Cancer Prev Res 2010; 3:108-13.
51. González-Sarrías A, Miguel V, Merino G i wsp. The gut microbiota ellagic acid-derived metabolite urolithin a and its sulfate conjugate are substrates for the drug efflux transporter breast cancer resistance protein (ABCG2/BCRP). J Agric Food Chem 2013; 61:4352-9.
52. Wang Y, Qiu Z, Zhou B i wsp. In vitro antiproliferative and antioxidant effects of urolithin A, the colonic metabolite of ellagic acid, on hepatocellular carcinomas HepG2 cells. Toxicol in Vitro 2015; 29:1107-15.
53. Espín JC, Larrosa M, García-Conesa MT. Biological significance of urolithins, the gut microbial ellagic acid-derived metabolites: The evidence so far. Evid Based Complemen Altern Med 2013.
54. Larrosa M, González-Sarrías A, García-Conesa MT i wsp. Urolithins, ellagic acid-derived metabolites produced by human colonic microflora, exhibit estrogenic and antiestrogenic activities. J Agric Food Chem 2006; 54:1611-20.
55. Li Z, Henning SM, Lee RP i wsp. Pomegranate extract induces ellagitannin metabolite formation and changes stool microbiota in healthy volunteers. Food Funct 2015; 6:2487-95.
56. Gimènez-Bastida JA, Truchado P, Larrosa M i wsp. Urolithins, ellagitannin metabolites produced by colon microbiota, inhibit Quorum Sensing in Yersinia enterocolitica: Phenotypic response and associated molecular changes. Food Chem 2012; 132:1465-74.
57. Myszka K, Czaczyk K. Mechanizm quorum sensing jako czynnik regulujący wirulencję bakterii Gram-ujemnych. Postępy Hig Med Dośw 2010; 64:582-9.
58. Verzelloni E, Pellacani C, Tgliazucchi i wsp. Antiglycative and neuroprotective activity of colon-derived polyphenol catabolites. Mol Nutr Food Res 2011; 55:S35-S43.
59. Sala R, Mena P, Savi M i wsp. Urolithins at physiological concentrations affect the levels of pro-inflammatory cytokines and growth factor in cultured cardiac cells in hyperglucidic conditions. J Funct Foods 2015; 15:97-105.
60. Kang I, Kim Y, Tomás-Barberán FA. Urolithin A, C, and D, but not iso-urolithin A and urolithin B, attenuate triglyceride accumulation in human cultures of adipocytes and hepatocytes. Mol Nutr Food Res 2016; 60(5):1129-38.
61. Yuan T, Ma H, Liu W i wsp. Pomegranate’s neuroprotective effects against Alzheimer’s disease are mediated by urolithins, its ellagitannin-gut microbial derived metabolites. ACS Chem Neurosci 2016; 7:26-33.